Cladistics
科学雑誌については、Cladistics(journal)を参照してください
。しばしば「分岐学の命名法」または「分岐学の用語」と呼ばれる系統発生の命名法については、系統発生の命名法を参照してください
分岐学(/ KのL ə D ɪ S T ɪ K S / ;から古代ギリシャ語 κλάδος(kládos) ‘分岐’)へのアプローチである生物学的分類する生物はグループ( “に分類されるクレード”)に基づきます最新の共通祖先の仮説。仮定された関係の証拠は、通常、共有派生形質(共有派生形質)です。 より遠いグループや祖先には存在しません。理論的には、共通の祖先とそのすべての子孫は、クレードの一部です。ただし、経験的な観点からは、共通の祖先は、性格状態を観察できる分類群の関係の分岐論的仮説に基づく推論です。重要なのは、すべての子孫が包括的な祖先の分岐群にとどまっていることです。たとえば、ワームまたは魚という用語が厳密な分岐の枠組み内で使用された場合、これらの用語には人間が含まれます。これらの用語の多くは、通常、分岐学の外で、たとえば「グレード」として、副系統的に使用されます。放射線は分岐によって新しいサブクレードを生成しますが、実際には性的交配は非常に密接に関連するグループを曖昧にする可能性が
分岐学の技術と命名法は、生物学以外の分野にも適用されてきました。(系統分類を参照して)
分岐学は現在、生物を分類するために最も一般的に使用されている方法です。
コンテンツ
1 歴史
2 方法論
3 文字状態の用語5 批判
6 問題
7 生物学以外の分野で
8 も参照してください
9 脚注と参考文献
10 参考文献
11 外部リンク
歴史

ヴィリー・ヘニッヒ1972

1920年のピーターチャルマーズミッチェル

ロバート・ジョン・ティラード
分岐解析と分類学の学校で使用された元の方法は、ドイツの昆虫学者 ウィリ・ヘニッヒの研究から派生しました。彼はそれを系統分類学(彼の1966年の本のタイトルでもあります)と呼びました。「分岐学」と「分岐群」という用語は、他の研究者によって広められました。本来の意味での分岐学は、系統発生分析で使用される特定の一連の方法を指しますが、現在では、フィールド全体を指すために使用されることも
現在、分岐法と呼ばれている方法は、早くも1901年に登場し、ピーター・チャルマーズ・ミッチェルによる鳥の研究 と、その後の1921年のロバート・ジョン・ティラード(昆虫の場合)およびW.ジマーマン(植物の場合)が登場しました。 )1943年に用語「クレードは」によって1958年に導入されたジュリアン・ハクスリーによって鋳造された後、ルシアン・クエノット1940年、「cladogenesis」1958年、「分岐学」によってアーサーカインとハリソン1960年、 1965年のエルンスト・マイヤーによる「クラディスト」(ヘニングの学校の支持者)、1966年の「分岐学」。ヘニングは、彼自身のアプローチを「系統発生学」と呼んだ。彼のオリジナル配合の時から1970年代の終わりまで、分岐学ではとのシステマティックに分析と哲学的アプローチとして出場表形分類学とは、いわゆる進化分類学。表形分類学をすることで、この時点で擁護した数値分類学者 ピーター・スニースとロバート・ソーカル、およびによって進化分類学エルンスト・マイヤー。
もともとは、本質的には、1950年に出版された本の中でWilli Hennigによって考案されましたが、分岐学は1966年に英語に翻訳されるまで繁栄しませんでした(Lewin1997)。今日、分岐学は、形態学的データから系統樹を推測するための最も一般的な方法です。
1990年代に、効果的なポリメラーゼ連鎖反応技術の開発により、生物の生化学的および分子遺伝学的特性への分岐法の適用が可能になり、系統発生学に利用できるデータの量が大幅に拡大しました。同時に、コンピュータが生物とその特性に関する大量のデータを処理することを可能にしたため、分岐学は進化生物学で急速に普及しました。
方法論
系統発生学と
クラドグラム 参照: 系統樹
で
分岐法は、各共有文字の状態変換を、グループ化の潜在的な証拠として解釈します。Synapomorphies(共有された派生文字の状態)はグループ化の証拠と見なされますが、symplesiomorphies (共有された祖先の文字状態)はそうではありません。分岐解析の結果は、系統発生関係の最良の仮説を表すと解釈されるクラドグラム–ツリー型の図(樹状図)です。従来、このようなクラドグラムは主に形態学的特性に基づいて生成され、元々は手作業で計算されていましたが、現在では系統発生分析で遺伝子シーケンシングデータと計算系統学が一般的に使用されており、多くの系統発生学者は節約基準を放棄して、より「洗練された」ものにしています。しかし、文字状態変換のそれほど倹約的ではない進化モデル。分岐学者は、これらのモデルが実際の経験的データセットからより「真の」または「正しい」結果を回復するという証拠がないため、これらのモデルは不当であると主張します。
すべてのクラドグラムは、特定の方法で分析された特定のデータセットに基づいています。データセットは、分子的、形態学的、動物行動学的および/またはその他の文字と、単系統であると推定される遺伝子、個体、集団、種、またはより大きな分類群である可能性のある運用分類群(OTU)のリストで構成される表です。したがって、すべて一緒に1つの大きなクレードを形成します。系統発生分析は、その分岐群内の分岐パターンを推測します。上記の仮定の違反は言うまでもなく、異なるデータセットと異なる方法は、しばしば異なるクラドグラムをもたらします。どちらが正しい可能性が高いかを示すことができるのは、科学的調査だけです。
たとえば、最近まで、次のようなクラドグラムは、カメ、トカゲ、ワニ、鳥の間の祖先関係の正確な表現として一般的に受け入れられていました。
▼ カメ
カメ 双弓類 ♦ 鱗竜類
トカゲ 主竜類
ワニ形上目
ワニ 恐竜 鳥
この系統発生の仮説が正しければ、▼の近くのブランチにあるカメと鳥の最後の共通の祖先は、 ♦の近くにあるトカゲと鳥の最後の共通の祖先よりも早く住んでいました。ただし、ほとんどの分子的証拠は、次のようなクラドグラムを生成します。
双弓類 ♦ 鱗竜類
トカゲ 主竜形類 ▼ カメ
カメ 主竜類
ワニ形上目
ワニ 恐竜 鳥
これが正確である場合、カメと鳥の最後の共通の祖先は、トカゲと鳥の最後の共通の祖先よりも遅く生きました。クラドグラムは、進化の歴史を説明するための2つの相互に排他的な仮説を示しているため、多くても1つが正しいものです。
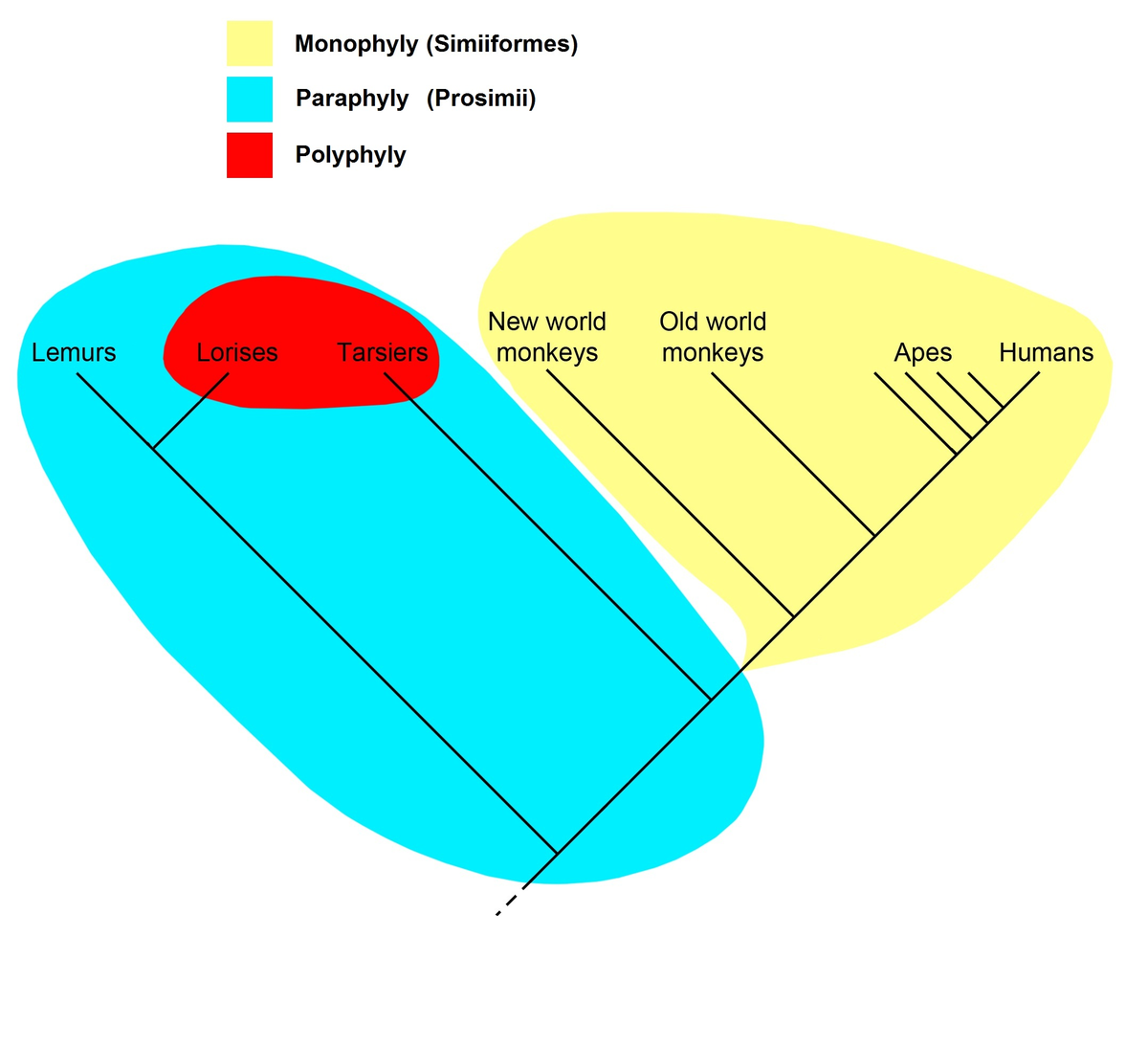
霊長類のクラドグラム
。単系統分類群( クレード:サルまたはAnthropoidea、黄色)、傍猿
分類
群(原猿、青色、赤いパッチを含む)、および
多系統分類群(夜間の霊長類– ロリスとtarsiers –赤)
右側のクラドグラムは、キツネザルやロリスなどの曲鼻亜目を含むすべての霊長類が共通の祖先を持ち、そのすべての子孫が霊長目である、または霊長目であったという現在の一般的に受け入れられている仮説を表しています。したがって、霊長類の名前はこの分岐群で認識されます。霊長類内では、すべての類人猿(サル、類人猿、および人間)は、すべての子孫が類人猿であるか、または類人猿であった共通の祖先を持っていたと仮定されているため、それらは類人猿と呼ばれる分岐群を形成します。一方、「原猿類」は側系統群を形成します。Prosimiiという名前は、クレードのみに名前を付ける系統分類では使用されません。代わりに、「原猿類」は、クレードStrepsirhiniとHaplorhiniに分けられ、後者にはメガネザル型下目と直鼻亜目が含まれます。
文字状態の用語
で
Hennigによって造られた次の用語は、グループ間で共有または別個の文字状態を識別するために使用されます。
共有原始形質( 「クローズフォーム」)または祖先の状態は、分類群がその祖先から保持しているキャラクターの状態です。互いに入れ子になっていない2つ以上の分類群が共有原始形質を共有する場合、それは共有原始形質です(syn-から「一緒に」)。共有原始形質は、その性格状態を示す分類群が必ずしも密接に関連していることを意味するものではありません。たとえば、爬虫類は伝統的に(とりわけ)冷血である(つまり、一定の高い体温を維持しない)ことを特徴としていますが、鳥は温血です。冷血は、伝統的な爬虫類や鳥の共通の祖先から受け継がれた共有原始形質であり、したがって、カメ、ヘビ、ワニ(とりわけ)の共有原始形質であるため、カメ、ヘビ、ワニが、鳥。
共有派生形質(「分離形式」)または派生状態は革新です。したがって、これを使用して、クレードを診断したり、系統分類でクレード名を定義したりすることができます。個々の分類群(特定の系統発生分析で単一の種または単一の末端によって表されるグループ)に由来する特徴は、固有派生形質(自動、「自己」から)と呼ばれます。固有派生形質は、グループ間の関係について何も表現し分岐群は、 (syn-から「一緒に」)共有派生形質によって識別(または定義)されます。たとえば、ホモ・サピエンスの指と相同な指の所持は、脊椎動物内の共有派生形質です。テトラポッドは、ホモ・サピエンスの数字と相同な最初の脊椎動物と、この脊椎動物のすべての子孫(共有派生形質に基づく系統学的定義)で構成されているものとして選択できます。重要なことに、数字を持たないヘビや他のテトラポッドはそれでもテトラポッドです。羊膜類の卵や双弓類の頭蓋骨などの他のキャラクターは、私たちと相同な数字を持っている祖先の子孫であることを示しています。
キャラクターの状態は、2つ以上の生物によって共有されているが、それらの共通の祖先または生物の1つにつながる系統の後続の祖先には存在しない場合、ホモプラスチックまたは「ホモプラシーのインスタンス」です。したがって、収束または逆転によって進化したと推測されます。哺乳類と鳥の両方が高い一定の体温を維持することができます(すなわち、彼らは温血動物です)。ただし、それらの重要な機能を説明する受け入れられたクラドグラムは、それらの共通の祖先がこの文字状態を欠くグループにあることを示しているため、状態は2つのクレードで独立して進化したに違いありません。温血動物は、哺乳類(またはより大きな分岐群)と鳥(またはより大きな分岐群)の共有派生形質ですが、これらの両方の分岐群を含むグループの共有派生形質ではありません。Hennigの補助原則は、共有されたキャラクターの状態は、他の証拠の重みと矛盾しない限り、グループ化の証拠と見なされるべきであると述べています。したがって、グループのメンバー間のいくつかの特徴のホモプラシーは、そのグループの系統発生仮説が確立された後にのみ推測される可能性が
共有原始形質と共有原始形質という用語は相対的です。それらのアプリケーションは、ツリー内のグループの位置によって異なります。たとえば、テトラポッドがクレードを形成するかどうかを判断しようとする場合、重要な問題は、4つの肢を持つことが、テトラポダに含まれる最も初期の分類群のシナポモルフィであるかどうかです。テトラポダの最も初期のメンバーはすべて、共通の祖先から4つの肢を継承しましたか。 、他のすべての脊椎動物はそうではなかった、または少なくとも相同ではなかったのか?対照的に、鳥などのテトラポッド内のグループの場合、4本の手足を持つことは共有原始形質です。これらの2つの用語を使用すると、相同性の議論の精度が高まり、特に、異なる相同性の特徴間の階層関係を明確に表現できます。
文字の状態が実際に同じであり、したがって単系統群を識別する可能性のある共有派生形質として分類できるかどうか、またはそれが同じように見えるだけであり、したがってそのようなものを識別できないホモプラシーであるかどうかを判断するのは難しい場合がありますグループ。循環論法の危険性が系統樹の形状に関する仮定は、文字の状態に関する決定を正当化するために使用され、その後、ツリーの形状の証拠として使用されます。 系統発生学は、そのような質問を決定するためにさまざまな形の倹約を使用します。到達する結論は、多くの場合、データセットとメソッドによって異なります。これが経験的科学の性質であり、このため、ほとんどの分岐学者はクラドグラムを関係の仮説と呼んでいます。多数のさまざまな種類の文字によってサポートされているクラドグラムは、より限定された証拠に基づくものよりも堅牢であると見なされます。
分類群の用語
単系統、傍系統、および多系統の分類群は、木の形状(上記のように)に基づいて、またそれらの特性の状態に基づいて理解することができます。 これらは以下の表で比較されています。
学期 ノードベースの定義 文字ベースの定義
単系統群 単系統分類群である分岐群は、推定された祖先のすべての子孫を含む分類群です。
分岐群は、1つ以上の共有派生形質によって特徴付けられます。つまり、分類群の最初のメンバーに存在し、その子孫に継承され(2番目に失われない限り)、他の分類群には継承されない派生文字状態です。
側系統群 側系統群は、クレードを取り、1つまたは複数の小さなクレードを取り除くことによって構築されるものです。(1つのクレードを削除すると、単一の側系統群が生成され、2つを削除すると、二重の側系統群が生成されます。)
側系統群は、1つまたは複数の共有原始形質によって特徴付けられます。つまり、祖先から継承されたが、すべての子孫に存在するわけではない文字の状態です。結果として、側系統群は、単系統分類群から1つまたは複数の分岐群を除外するという点で切り捨てられます。別名は進化グレードで、グループ内の祖先のキャラクターの状態を指します。側系統群は古生物学者や進化分類学者の間で人気がありますが、分岐群は正式な情報コンテンツを持っているとは認識しそれらは単なる分岐群の一部です。
多系統 多系統群は、単系統でも側系統でもない群集です。 多系統群は、1つまたは複数のホモプラシーによって特徴付けられます。つまり、同じになるように収束または復帰したが、共通の祖先から継承されていない文字の状態です。生態学者は、多系統群集を生態学的コミュニティの機能的参加者(例えば、一次生産者、腐食生物など)にとって意味のあるラベルと見なすことがありますが、系統分類学者は多系統群集を分類学的に意味のある実体として認識しません。
批判
分岐学は、一般的または特定のアプリケーションのいずれかで、最初から批判されてきました。特定の性格状態が相同であるかどうかに関する決定は、それらが共有派生形質であるという前提条件であり、循環論法と主観的判断を伴うものとして異議を唱えられてきました。もちろん、証拠の潜在的な信頼性の欠如は、体系的な方法、さらに言えば、経験的な科学的努力にとっては問題です。
変換された分岐学は、分岐学分析から系統学に関する先験的な仮定を取り除くことによってこれらの問題のいくつかを解決しようとして1970年代後半に発生しましたが、それは人気がないままです。
問題
分岐法では、化石種を分岐群の実際の祖先として識別しません。代わりに、化石分類群は別々の絶滅した枝に属するものとして識別されます。化石種は実際の分岐群の祖先である可能性がありますが、それを知る方法はありません。したがって、より保守的な仮説は、共有派生形質のパターンによって暗示されるように、化石分類群が他の化石および現存する分類群に関連しているというものです。
生物学以外の分野で
クラドグラムの基礎となるデータを取得するために使用される比較は、生物学の分野に限定されません。共通の祖先を持っていると仮定され、共通の特性のセットが適用される場合と適用されない場合がある個人またはクラスのグループは、ペアごとに比較できます。クラドグラムは、多くの異なる学問領域のアイテムのグループ内の仮想的な降下関係を表すために使用できます。唯一の要件は、アイテムが識別および測定できる特性を備えていることです。
人類学と考古学:分岐法は、文化的特徴または人工物の特徴のグループを使用して、文化または人工物の発達を再構築するために使用されてきました。
比較神話学と民話は、多くの神話の原型を再構築するために分岐法を使用しています。神話素で構築された神話の系統発生は、低い水平感染(借用)、歴史的な(旧石器時代の)拡散、断続平衡説を明確にサポートしています。それらはまた、民話間の異文化間の関係についての仮説をテストするための強力な方法です。
文学:カンタベリー物語の生き残った写本とサンスクリット語のチャラカ・サンヒタの写本の分類には、分岐法が使用されてきました。
歴史言語学:分岐法は、言語学的特徴を使用して言語の系統発生を再構築するために使用されてきました。これは、歴史言語学の従来の比較方法に似ていますが、節約の使用においてより明確であり、大規模なデータセットのはるかに高速な分析を可能にします(計算系統学)。
本文批評または語幹論: 分岐法は、同じ作品の写本の系統発生を再構築する(そして失われたオリジナルを再構築する)ために、独特のコピーエラーを共有派生形質として使用してきました。これは、編集者が手動で処理することが不可能な多数の変種を含む多数の原稿のグループを評価し、遺伝的関係に置くことができるという点で、従来の歴史比較言語学とは異なります。また、妥当な期間内に手動で評価することは不可能な、汚染された感染の伝統の節約分析も可能にします。
天体物理学は、銀河間の関係の歴史を推測して、銀河多様化の分岐図仮説を作成します。
も参照してください
バイオインフォマティクス生物数学 合体理論 共通祖先
科学的命名の用語集
語族
パトロクラドグラム
系統発生ネットワーク
科学的分類 Stratocladistics サブクレード
分類学
3分類群分析
ツリーモデル
木の構造
![]()
生物学ポータル
![]()
進化生物学ポータル
脚注と参考文献
^ ハーパー、ダグラス。「クレード」。オンライン語源辞書。
^ コロンビア百科事典 ^ 「分岐学入門」。Ucmp.berkeley.edu 。
^ オックスフォード英語辞典 ^ オックスフォード英語辞典 ^ 「分岐学の必要性」。www.ucmp.berkeley.edu 。
^ Brinkman&Leipe 2001、p。323 ^ シュー、ランドール。2000. Biological Systematics:Principles and Applications、p.7(Nelson and Platnick、1981を引用)。コーネル大学出版局(books.google)
^ Folinsbee、Kaila etal。2007. 5系統発生学への定量的アプローチ、p。172.メックス牧師 Div。225-52(kfolinsb.public.iastate.edu)
^ Craw、RC(1992)。「分岐学のマージン:系統分類学の出現におけるアイデンティティ、違いおよび場所」。グリフィスでは、PE(編)。生命の木:生物学の哲学のエッセイ。ドルドレヒト:クルーワーアカデミック。pp。65–107。ISBN 978-94-015-8038-0。
^ シュー、ランドール。2000.生物学的分類学:原理と応用、p.7。Cornell U. Press ^ Cuénot1940 ^ Webster の9番目の新しい大学辞書 ^ Cain&Harrison 1960 ^ Dupuis 1984 ^ Weygoldt 1998 ^ リンダル、エイリック; Brower、Andrew VZ(2011)、「モデルベースの系統発生分析は節約よりも優れていますか?経験的データを使用したテスト」、Cladistics、27(3):331–334、doi:10.1111 /j.1096-0031.2010.00342。 x、S2CID 84907350 ^ Jerison 2003、p。254 ^ Benton、Michael J.(2005)、Vertebrate Palaeontology、Blackwell、pp.214、233、ISBN 978-0-632-05637-8 ^ ライソン、タイラー; ギルバート、スコットF.(2009年3月から4月)、「カメはずっと下にある:ケロニアの木の根元にあるロガーヘッド」(PDF)、Evolution&Development、11(2):133–135、CiteSeerX10.1.1.695 。 4249、doi:10.1111 / j.1525-142X.2009.00325.x、PMID 19245543、S2CID 3121166
^ Patterson 1982、pp。21–74 ^ Patterson 1988
^ Pinna 1991 ^ ローリン&アンダーソン2004 ^ Hennig 1966 ^ James&Pourtless IV 2009、p。25:「共有派生形質は仮説を守るために呼び出されます。仮説は共有派生形質を守るために呼び出されます。」 ^ Brower、AVZおよびSchuh、RT。2021.生物学的分類学:原理と応用(第3版)。Cornell University Press、ニューヨーク州イサカ ^ パターソン1982 ^ 多くの情報源は、欠落しているグループが単系統である必要のない「側系統群」の口頭での定義を示しています。ただし、側系統群を表す図が表示される場合、これらは常に欠落しているグループを単系統として示します。たとえば、Wiley etal。を参照して1991年、p。4 ^ テイラー2003 ^ Adrain、Edgecombe&Lieberman 2002、pp。56–57 ^ Oreskes、Naomi、Kristin Shrader-Frechette、およびKennethBelitz。「地球科学における数値モデルの検証、妥当性確認、および確認。」科学263、いいえ。5147(1994):641-646。
^ NilsMøllerAnderson、2001年W.ヘニッヒの「系統分類学」が現代の昆虫学に与える影響。J.Entomol。98:133-150 オンライン
^ プラトニック、ノーマンI.「哲学と分岐学の変容」。系統的動物学28、いいえ。4(1979):537-546 ^ ブロワー、アンドリューVZ。「50色のクラディズム。」生物学と哲学33、いいえ。1-2(2018):8。
^ クレル、フランク-T; クランストン、ピーターS.(2004)。「木のどちら側がより基本的ですか?:社説」。体系的な昆虫学。29(3):279–281。土井:10.1111 /j.0307-6970.2004.00262.x。S2CID82371239。_ ^ パターソン、コリン。「進化の関係を決定する上での化石の重要性。」生態学と分類学の年次レビュー12、no。1(1981):195-223 ^ Mace、Clare&Shennan 2005、p。1 ^ Lipo etal。2006年 ^ d’Huy 2012a、b; d’Huy 2013a、b、c、d ^ ロスと他。2013年 ^ Tehrani 2013 ^ 「カンタベリー物語プロジェクト」。
^ Maas 2010–2011 ^ Oppenheimer 2006、pp。290–300、340–56 ^ ロビンソン&オハラ1996 ^ Fraix-Burnet etal。2006年
参考文献
アドレーン、ジョナサンM。; Edgecombe、Gregory D.&Lieberman、Bruce S.(2002)、Fossils、Phylogeny、and Form:An Analytical Approach、New York:Kluwer Academic、ISBN 978-0-306-46721-9、
Baron、C。&Høeg、JT(2005)、 “”Gould、Scharm and the paleontologocalspective in Evolutionary biology””、in Koenemann、S。&Jenner、RA(eds。)、Crustacea and Arthropod Relationships、CRC Press、 pp。3 –14、ISBN 978-0-8493-3498-6、
ベントン、MJ(2000)、「ステム、ノード、クラウンクレード、およびランクフリーリスト:リンネは死んでいますか?」 (PDF)、Biological Reviews、75(4):633–648、CiteSeerX 10.1.1.573.4518、doi:10.1111 / j.1469-185X.2000.tb00055.x、PMID 11117201、S2CID 17851383 、オリジナル からアーカイブ( PDF) 2017年8月9日、
ベントン、MJ(2004)、脊椎動物古生物学(第3版)、オックスフォード:ブラックウェルサイエンス、ISBN 978-0-632-05637-8
Brinkman、Fiona SL&Leipe、Detlef D.(2001)、””Phylogenetic analysis”” (PDF)、in Baxevanis、Andreas D.&Ouellette、BF Francis(eds。)、Bioinformatics:実用的なガイド。 (第2版)、pp。323–358、2013年10月20日にオリジナル (PDF)からアーカイブ、
カイン、AJ; ジョージア州ハリソン(1960)、「Phyletic weighting」、Proceedings of the Zoological Society of London、35:1–31
Cuénot、Lucien(1940)、「Remarques sur unessaid’arbregénéalogiquedurègneanimal」、Comptes Rendusdel’AcadémiedesSciencesdeParis、210:23–27。Gallicaでオンラインで無料で入手できます(直接URLはありません)。これは、「クレード」という用語の最初の使用について、Hennig1979によってクレジットされた論文です。
Dupuis、Claude(1984)、「Willi Hennigの分類学的思考への影響」、Annual Review of Ecology and Systematics、15:1–24、doi : 10.1146 / annurev.ecolsys.15.1.1、ISSN0066-4162 。
Farris、James S.(1977)、「脊椎動物分類への表形的アプローチについて」、MK、Hecht; グッディ、PC; Hecht、BM(eds。)、脊椎動物の進化における主要なパターン、プレナム、ニューヨーク、pp。823–850
Farris、James S.(1979a)、「系統分類の自然性について」、Systematic Zoology、28(2):200–214、doi:10.2307 / 2412523、JSTOR 2412523
Farris、James S.(1979b)、「系統発生システムの情報内容」、Systematic Zoology、28(4):483–519、doi:10.2307 / 2412562、JSTOR 2412562
Farris、James S.(1980)、「系統発生システムの効率的な診断」、Systematic Zoology、29(4):386–401、doi:10.2307 / 2992344、JSTOR 2992344
Farris、James S.(1983)、「系統発生分析の論理的基礎」、Platnick、Norman I。; Funk、Vicki A.(eds。)、Advances in Cladistics、vol。2、コロンビア大学出版、ニューヨーク、7〜36ページ
Fraix-Burnet、D。; Choler、P。; Douzery、EJP; Verhamme、A。(2006)、「Astrocladistics:A Phylogenetic Analysis of Galaxy EvolutionII。FormationandDiversification of Galaxys」、Journal of Classification、23(1):57–78、arXiv:astro-ph / 0602580、Bibcode:2006JClas ..23 … 57F、doi:10.1007 / s00357-006-0004-4、S2CID 2537194
Hennig、Willi(1966)、系統分類学(tr。D.Dwight Davis and Rainer Zangerl)、イリノイ州アーバナ:大学 イリノイ大学出版局(1979年と1999年に転載)、ISBN 978-0-252-06814-0
Hennig、Willi(1975)、「’分岐解析または分岐分類?’:Ernst Mayrへの回答」 (PDF)、系統動物学、24(2):244–256、doi:10.2307 / 2412765、JSTOR 2412765、1974年5月に対応。
Hennig、Willi(1999)、系統分類学(1966年の本の第3版)、Urbana:University of Illinois Press、ISBN 978-0-252-06814-0 最終的に1982年に出版されたドイツ語の原稿から翻訳された(Phylogenetische Systematik、Verlag Paul Parey、ベルリン)。
Hull、David(1988)、Science as a Process、シカゴ大学出版局、ISBN 978-0-226-36051-5
James、Frances C.&Pourtless IV、John A.(2009)、Cladistics and the Origin of Birds:A Review and Two New Analyses (PDF)、Ornithological Monographs、No。66、American Ornithologists’Union、ISBN 978-0-943610-85-6、
d’Huy、Julien(2012a)。「Unoursdanslesétoiles:recherchephylogénétiquesurunmythepréhistorique」。PréhistoireduSud-Ouest。20(1):91–106。
d’Huy、Julien(2012b)、「Lemotif de Pygmalion:origine afrasienne etdiffuseenAfrique」。サハラ、23:49-59 。
d’Huy、Julien(2013a)、「Polyphemus(Aa。Th.1137)」。「先史時代の物語の系統発生的再構築」。NouvelleMythologieComparée/新しい比較神話1、
d’Huy、Julien(2013b)。「神話の系統学的アプローチとその考古学的結果」。ロックアートリサーチ。30(1):115–118。
d’Huy、Julien(2013c)「Lesmythesévolueraientparponctuations」。Mythologiefrançaise、252、2013c:8-12。
d’Huy、Julien(2013d)「ベルベル人の空での宇宙狩り:旧石器時代の神話の系統発生的再構築」。Les Cahiers de l’AARS、15、2013d:93-106。
ジェリソン、ハリーJ.(2003)、「比較心理学の理論について」、スタンバーグ、ロバートJ。; カウフマン、ジェームズC.(編)、インテリジェンスの進化、ニュージャージー州マフア:ローレンスエルバウムアソシエイツ社、ISBN 978-0-12-385250-2
Laurin、M。&Anderson、J。(2004)、「科学文献における テトラポダという名前の意味:交換」(PDF)、系統生物学、53(1):68–80、doi:10.1080 / 10635150490264716、PMID 14965901
リポ、カール; オブライエン、マイケルJ。; コラード、マーク; et al。、eds。(2006)、私たちの祖先のマッピング:人類学と先史時代の系統発生的アプローチ、ピスカタウェイ:トランザクションパブリッシャー、ISBN 978-0-202-30751-0
Maas、Philipp(2010–2011)、Jürgen、Hanneder; Maas、Philipp(eds。)、 “”Computer Aided Stemmatics – The Case of 52 Text VersionsofCarakasasaṃhitāVimānasthāna8.67-157″”、WienerZeitschriftfürdieKundeSüdasiens、52–53:63–120、doi:10.1553 / wzks2009- 2010s63
メイス、ルース; クレア、クレアJ。; シェナン、スティーブン、編 (2005)、文化的多様性の進化:系統発生的アプローチ、ポートランド:Cavendish Press、ISBN 978-1-84472-099-6
Mayr、Ernst(1974)、「分岐解析または分岐分類?」 (PDF)、ZeitschriftfürZoologischeSystematikund Evolutionsforschung、12:94–128、doi:10.1111 / j.1439-0469.1974.tb00160.x
Mayr、Ernst(1976)、Evolution and thediversity of life(Selected essays)、Cambridge、Massachusetts:Harvard University Press、ISBN 978-0-674-27105-01997年にペーパーバックで再発行されました。Mayrの1974年の反分岐学論文のpp。433–476、「分岐解析または分岐分類」の再版が含まれています。これは、Hennig1975が回答した論文です。
マイヤー、エルンスト(1978)、 “”起源と体系的かつ進化生物学におけるいくつかの用語の歴史””、体系的動物学、27(1):83-88、DOI:10.2307 / 2412818、JSTOR 2412818。
Mayr、Ernst(1982)、生物学的思考の成長:多様性、進化、継承、マサチューセッツ州ケンブリッジ:ハーバード大学出版局、ISBN 978-0-674-36446-2
オッペンハイマー、スティーブン(2006)、英国の起源、ロンドン:ロビンソン、ISBN 978-0-7867-1890-0
パターソン、コリン(1982)、「形態学的特徴と相同性」、ジョイジー、ケネスA; 金曜日、AE(eds。)、系統発生的再構築の問題、系統分類学協会特別巻21、ロンドン:アカデミックプレス、ISBN 978-0-12-391250-3。
Patterson、Colin(1988)、「古典的および分子生物学における相同性」、Molecular Biology and Evolution、5(6):603–625、doi:10.1093 / oxfordjournals.molbev.a040523、PMID 3065587
de Pinna、MGG(1991)、「 分岐パラダイムにおける相同性の概念とテスト」(PDF)、Cladistics、7(4):367–394、CiteSeerX 10.1.1.487.2259、doi:10.1111 / j.1096-0031.1991 .tb00045.x、S2CID 3551391、2011年7月22日にオリジナル (PDF)からアーカイブ、
デ・ケイロス、K.&ゴーティエ、J.(1992)、””系統発生分類”” (PDF) 、エコロジーと分類学の年次レビュー、23:449から480、DOI:10.1146 / annurev.ecolsys.23.1.449からアーカイブ、オリジナル (PDF) 2012年3月20日には、検索された7月28日の2012
Robinson、Peter MW&O’Hara、Robert J.(1996)、「古ノルド語写本の伝統の古典的分析」、Research in Humanities Computing、4:115–137 、2010年12月13日検索
ロス、ロバートM。; グリーンヒル、サイモンJ。; Atkinson、Quentin D.(2013)。「ヨーロッパの民話の人口構造と文化地理学」。王立協会紀要B:生物科学。280(の1756):20123065. DOI:10.1098 / rspb.2012.3065。PMC 3574383。PMID 23390109。
Schuh、Randall T.&Brower、Andrew VZ(2009)、Biological Systematics:Principles and Applications(2nd ed。)、Cornell University Press、ISBN 978-0-8014-4799-0
Taylor、Mike(2003)、単系統、側系統、多系統などの用語はどういう意味ですか?、
Tehrani、Jamshid J.、2013年、「赤ずきんの系統発生」、PLOS ONE、11月13日。
Tremblay、Frederic(2013)、””Nicolai Hartmann and the Metaphysical Foundation of Phylogenetic Systematics””、Biological Theory、7(1):56–68、doi:10.1007 / s13752-012-0077-8、S2CID 84932063
Weygoldt、P。(1998年2月)、「鋏角類の進化と分類学」、実験および応用ダニ学、22(2):63–79、doi:10.1023 / A:1006037525704、S2CID 35595726
Wheeler、Quentin(2000)、種の概念と系統発生理論:討論、コロンビア大学出版、ISBN 978-0-231-10143-1
ワイリー、EO; Siegel-Causey、D。; Brooks、DR&Funk、VA(1991)、「第1章はじめに、用語と概念」、The Compleat Cladist:A Primer of Phylogenetic Procedures (PDF)、カンザス大学自然史博物館、ISBN 978-0-89338-035-9、からアーカイブ元 (PDF) 2010年12月3日に、検索された12月13日と2010年
ペンシルベニア州ウィリアムズ(1992)、「クラディズムの混乱」、Synthese、01(1–2):135–152、doi:10.1007 / BF00484973、S2CID 46974048
外部リンク
を聞く(13分)
![]()
このオーディオファイルは、2005年4月30日付けのの改訂版から作成されたもの
であり、その後の編集は反映され
(2005-04-30)( 音声ヘルプ ・
より多くの話された記事)
コモンズの分岐学に関連するメディア
OneZoom:Tree of Life –直感的でズーム可能なフラクタルエクスプローラーとしてのすべての生物(レスポンシブデザイン)
ヴィリー・ヘニッヒ協会
Cladistics(Willi Hennig Societyの学術雑誌)
コリンズ、アレンG。; Guralnick、Rob; スミス、デイブ(1994–2005)。「系統分類学への旅」。カリフォルニア大学古生物学博物館。
フェルゼンシュタイン、ジョー。「系統発生プログラム」。シアトル:ワシントン大学。
オニール、デニス(1998–2008)。「生物の分類」。カリフォルニア州サンマルコス:パロマー大学。
ロビンソン、ピーター; オハラ、ロバートJ.(1992)。「1991年のテキスト批評チャレンジに関するレポート」。rjohara.net 。
テオバルド、ダグラス(1999–2004)。「系統発生入門書」。TalkOriginsアーカイブ。”