DD-transpeptidase
DD-トランスペプチダーゼ(EC 3.4.16.4、 DD-ペプチダーゼ、 DD-トランスペプチダーゼ、 DD-カルボキシペプチダーゼ、D-アラニル-D-アラニンカルボキシペプチダーゼ、 D-アラニル-D-アラニン切断-ペプチダーゼ、D-アラニンカルボキシペプチダーゼカルボキシペプチダーゼ、およびセリン型D-Ala-D-Alaカルボキシペプチダーゼ。 )は、RL-aca- D-アラニル-D-アラニンカルボニルのRL-aca- D-アラニル部分の転移を触媒する細菌酵素です。それらの活性部位セリンのγ-OHへのドナーおよびこれから最終アクセプターへのドナー。これは、細菌の細胞壁生合成、すなわちペプチドグリカン鎖のペプチド側鎖を架橋するペプチド転移に関与しています。
セリン型D-Ala-D-Alaカルボキシペプチダーゼ
ストレプトマイセスK15DD-トランスペプチダーゼの構造
識別子
EC番号
3.4.16.4
CAS番号
9077-67-2
データベース IntEnz IntEnzビュー
ブレンダ
BRENDAエントリー ExPASy NiceZymeビュー KEGG KEGGエントリー MetaCyc 代謝経路
プリアモス
プロフィール
PDB構造
RCSB PDB PDBe PDBsum索 PMC
記事 PubMed 記事 NCBI タンパク質
抗生物質 ペニシリンは、非常に安定したペニシロイル酵素中間体を形成することにより、トランスペプチダーゼ酵素に不可逆的に結合し、その活性を阻害します。ペニシリンとトランスペプチダーゼの間の相互作用のため、この酵素はペニシリン結合タンパク質(PBP)としても知られています。
コンテンツ
1 機構
2 構造
3 生物学的機能
4 病気の関連性
5 も参照してください
6 参考文献
7 外部リンク
機構
DD-トランスペプチダーゼは、トリプシンタンパク質ファミリーのタンパク質分解反応と機構的に類似しています。
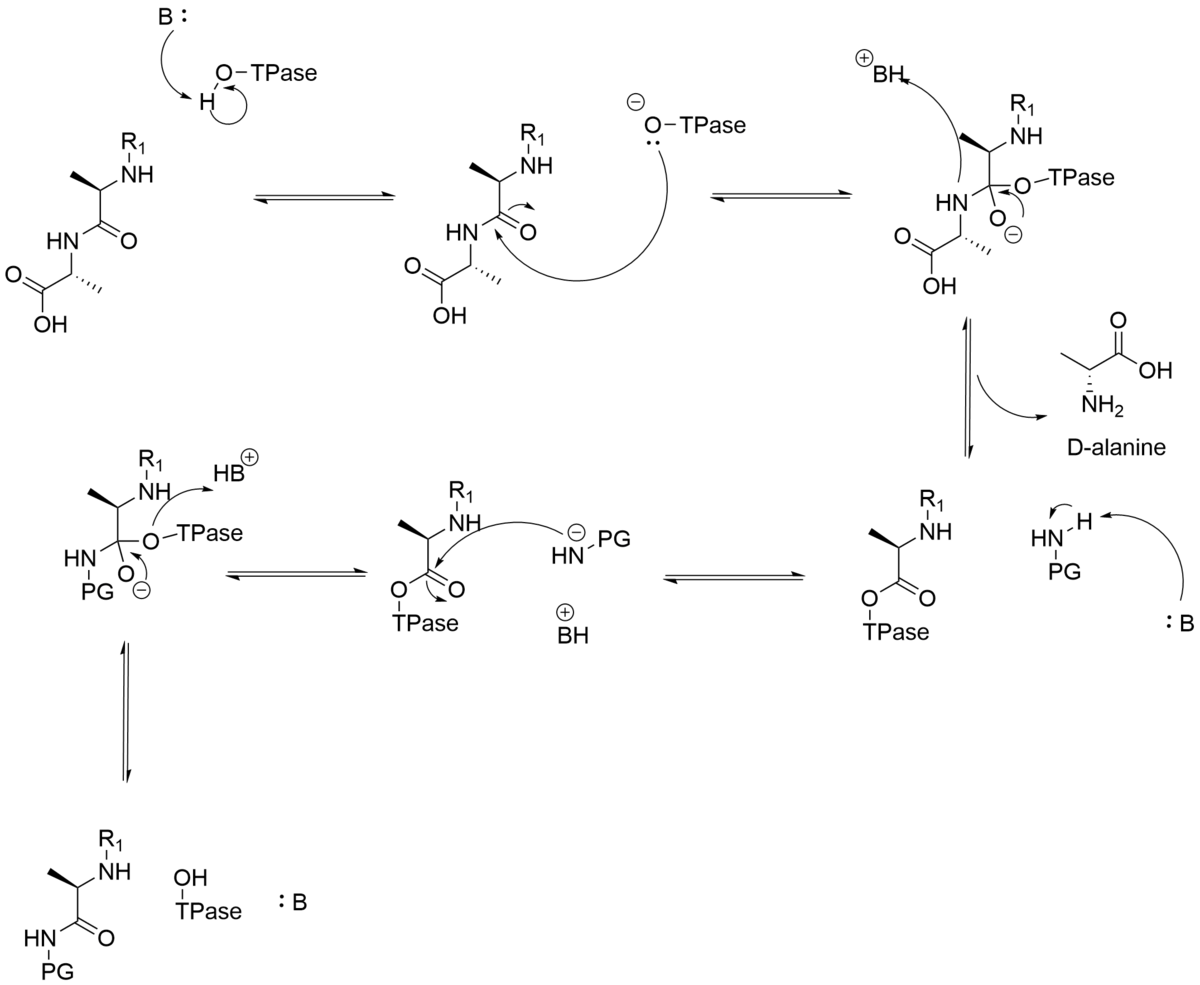
DD-トランスペプチダーゼの触媒機構
隣接するグリカン鎖のペプチジル部分の架橋は、2段階の反応です。最初のステップは、カルボニル供与体として作用するペプチドユニット前駆体のD-アラニル-D-アラニン結合の切断、カルボキシル末端D-アラニンの放出、およびアシル酵素の形成を含む。2番目のステップでは、アシル酵素中間体を分解し、D-アラニル部分のカルボニルと別のペプチドユニットのアミノ基との間に新しいペプチド結合を形成します。
DD-ペプチダーゼメカニズムのほとんどの議論は、プロトン移動の触媒を中心に展開しています。アシル酵素中間体の形成中に、プロトンを活性部位のセリンヒドロキシル基から除去し、アミン脱離基にプロトンを付加する必要が同様のプロトンの動きは、脱アシル化において促進されなければなりません。これらのプロトン移動に関与する一般的な酸および塩基触媒の正体はまだ解明されしかしながら、触媒トライアドのチロシン、リジン、およびセリン、ならびにセリン、リジン、セリンが提案されている。
構造
トランスペプチダーゼは、ペニシロイル-セリントランスフェラーゼスーパーファミリーのメンバーであり、 SxxKで保存された特徴的なモチーフを持っています。「x」は可変アミノ酸残基を示し、このスーパーファミリーのトランスペプチダーゼは、SxxK、SxN(またはアナログ)、およびKTG(またはアナログ)の3つのモチーフの形で傾向を示します。これらのモチーフは同等の場所にあり、ポリペプチド鎖に沿ってほぼ等間隔に配置されています。折りたたまれたタンパク質は、これらのモチーフを、 all-αドメインとα/βドメインの間の触媒中心で互いに近づけます。
ストレプトマイセスK15DD-トランスペプチダーゼの構造が研究されており、2つのドメインに組織化された単一のポリペプチド鎖で構成されています。1つのドメインは主にα-ヘリックスを含み、2番目のドメインはα/β-タイプです。触媒裂の中心は、Ser35-Thr36-Thr37-Lys38テトラッドで占められています。これには、ヘリックスα2のアミノ末端に求核性のSer35残基が含まれています。空洞の片側は、らせんα4とα5を接続するSer96-Gly97-Cys98ループによって定義されます。Lys213-Thr214-Gly215トライアドは、空洞の反対側のストランドβ3上に必須のSer35残基のバックボーンNH基とモチーフLys213-Thr214-Gly215の下流のSer216のバックボーンNH基は、触媒作用に必要なオキシアニオンホール機能と互換性のある位置を占めています。
カルボニル供与体の感受性ペプチド結合がD-配置を持つ2つの炭素原子の間に伸びているため、この酵素はDD-トランスペプチダーゼとして分類されます。
生物学的機能
すべての細菌は、少なくとも1つ、ほとんどの場合はいくつかの単機能セリンDDペプチダーゼを持っています。
病気の関連性

(A)ペプチドグリカン末端のD-Ala-D-Ala末端と(B)ペニシリンの間の構造的類似性。トランスペプチダーゼは、TPase触媒反応に対してペニシリンを誤認識します。
この酵素は、必須であり、ペリプラズムからアクセス可能であり、哺乳動物細胞には同等のものがないため、優れた創薬ターゲットです。DD-トランスペプチダーゼは、 β-ラクタム系抗生物質(ペニシリンなど)の標的タンパク質です。これは、β-ラクタムの構造がD-ala-D-ala残基に非常に似ているためです。
β-ラクタムは、セリンDD-トランスペプチダーゼ触媒部位を競合的に不活性化することによってその効果を発揮します。ペニシリンは、D-Ala-D-Ala末端カルボニル供与体の環状類似体であるため、この抗生物質の存在下では、セリンエステル結合ペニシロイル酵素のレベルで反応が停止します。したがって、β-ラクタム系抗生物質は、これらの酵素をペニシリン結合タンパク質のように振る舞わせます。
速度論的には、DD-ペプチダーゼとベータラクタム間の相互作用は3段階の反応です。E + I ⇌ E ⋅ IE − I ∗ E + P { E + I rightleftharpoons E cdot I rightarrow EI * rightarrow E + P}
ベータ-イアクタムは、DD-トランスペプチダーゼと高い安定性の付加物EI *を形成する可能性がこの付加物の半減期は約数時間ですが、通常の反応の半減期はミリ秒のオーダーです。
細胞壁形成の原因となる酵素プロセスへの干渉は、細菌の自己消化システムの誘発により、細胞の溶解と死をもたらします。
も参照してください
バンコマイシン、D-ala-D-ala残基に結合し、グリコシルトランスフェラーゼを介して伸長を阻害する抗生物質
参考文献
^ 「EC3.4.16.4セリン型D-Ala-D-Alaカルボキシペプチダーゼ」。酵素構造データベース。2006年5月17日にオリジナルからアーカイブされました。
^ Grandchamps J、Nguyen-DistècheM、Damblon C、FrèreJM、Ghuysen JM(1995)。「ストレプトマイセスK15活性部位セリンDD-トランスペプチダーゼ:ペプチド、チオールエステルおよびエステルカルボニルドナーの特異性プロファイルと転移反応の経路」。BiochemJ。307(Pt 2)(2):335–9。土井:10.1042 / bj3070335。PMC1136653。_ PMID7733866。_
^ Yocum RR、Waxman DJ、Rasmussen JR、Strominger JL(1979)。「ペニシリン作用のメカニズム:ペニシリンと基質は、2つの細菌性D-アラニンカルボキシペプチダーゼの同じ活性部位セリンに共有結合します」。Proc Natl Acad SciUSA。76(6):2730–4。Bibcode:1979PNAS … 76.2730Y。土井:10.1073 /pnas.76.6.2730。PMC383682。_ PMID111240。_
^ Gordon E、Mouz N、DuéeE、Dideberg O。「Streptococcuspneumoniae由来のペニシリン結合タンパク質2xの結晶構造とそのアシル酵素型:薬剤耐性への影響」。分子生物学ジャーナル。299(2):477–85。土井:10.1006 /jmbi.2000.3740。PMID10860753。_
^ Goffin C、Ghuysen JM。「SxxKスーパーファミリーアシルトランスフェラーゼの生化学および比較ゲノミクスは、マイコバクテリアのパラドックスへの手がかりを提供します:ペニシリン感受性標的タンパク質の存在対治療薬としてのペニシリンの効率の欠如」。微生物学および分子生物学のレビュー。66(4):702–38、目次。土井:10.1128 /MMBR.66.4.702-738.2002。PMC134655。_ PMID12456788。_
^ dFonzéE 、Vermeire M、Nguyen-DistècheM、Brasseur R、Charlier P(1999年7月)。「中程度のペニシリン感受性のペニシロイル-セリントランスフェラーゼの結晶構造。ストレプトマイセスK15のDD-トランスペプチダーゼ」。Journal of BiologicalChemistry。274(31):21853–60。土井:10.1074 /jbc.274.31.21853。PMID10419503。_
^ プラットRF。「細菌のDD-ペプチダーゼ(ペニシリン結合タンパク質)の基質特異性」。細胞および分子生命科学。65(14):2138–55。土井:10.1007 / s00018-008-7591-7。PMID18408890。_ S2CID25147733。_
^ Walsh C、Wencewicz T(2016)。抗生物質:課題、メカニズム、機会(第2版)。アメリカ微生物学会(Verlag)。ISBN
978-1-55581-930-9。
^ Ghuysen JM(1994年10月)。「ペニシリン結合タンパク質とベータラクタマーゼの分子構造」。微生物学の動向。2(10):372–80。土井:10.1016 / 0966-842X(94)90614-9。PMID7850204。_
^ Kelly JA、Kuzin AP、Charlier P、FonzéE(1998年4月)。「ペニシリンと相互作用する酵素のX線研究」。細胞および分子生命科学(投稿原稿)。54(4):353–8。土井:10.1007 / s000180050163。hdl:2268/77968。PMID9614972。_ S2CID21504958。_
^ Nguyen-DistècheM、Leyh-Bouille M、Ghuysen JM(1982年10月)。「ペニシリン感受性D-アラニル-D-アラニン切断トランスペプチダーゼの形でのストレプトマイセスK15株の膜結合26000-Mrペニシリン結合タンパク質の単離」。生化学ジャーナル。207(1):109–15。土井:10.1042 / bj2070109。PMC1153830。_ PMID7181854。_
^ Ghuysen JM、FrèreJM、Leyh-Bouille M、Nguyen-DistècheM、Coyette J、Dusart J、Joris B、Duez C、Dideberg O、Charlier P(1984)。「細菌壁ペプチドグリカン、DD-ペプチダーゼおよびベータラクタム抗生物質」。感染症のスカンジナビアジャーナル。補足。42:17–37。PMID6597561。_
^ Spratt BG(1983年5月)。「ペニシリン結合タンパク質とベータラクタム抗生物質の将来。第7回フレミング講義」。一般微生物学ジャーナル。129(5):1247–60。土井:10.1099 / 00221287-129-5-1247。PMID6352855。_
外部リンク
ペプチダーゼとその阻害剤のMEROPSオンラインデータベース: S11.001
EC 3.4.16.4
米国国立医学図書館の医学主題見出し(MeSH)のセリンタイプ+ D-アラ-D-アラ+カルボキシペプチダーゼ
ポータル:
![]()
生物学”