List_of_African_dinosaurs
これは、アフリカから遺体が回収された恐竜のリストです。アフリカには豊富な化石記録がありますが、それはまだらで不完全です。三畳紀と前期ジュラ紀の恐竜が豊富です。これらの時代のアフリカの恐竜には、コエロフィシス、ドラコヴェナトル、メラノロサウルス、マッソスポンディルス、ユースケロサウルス、ヘテロドントサウルス、アブリクトサウルス、レソトサウルスが含まれます。中期ジュラ紀では、竜脚類のアトラサウルス、シェブサウルス、ジョバリア、スピノフォロサウルス、そして獣脚類のアフロベナトルが栄えました。後期ジュラ紀は、主にタンザニアのリンディ地域の壮大なテンダグル層のおかげで、アフリカでよく表されています。ヴェテルプリスティサウルス、オスタフリカサウルス、エラフロサウルス、ギラッファティタン、ディクラエオサウルス、ジャネンシア、トルニエリア、テンダグリア、ケントロサウルス、ディサロトサウルスは、テンダグリアから遺体が回収された恐竜の1つです。この動物相は、米国のモリソン累層やポルトガルのロウリニャン層と強い類似性を示しているようたとえば、同様の獣脚類、鳥脚類、竜脚類がテンダグル層とモリソン層の両方で発見されています。これは、生物地理学的に重要な意味を持っています。
アフリカの前期白亜紀は、主に大陸の北部、特にニジェールから知られています。スコミムス、エルハゾサウルス、レッバキサウルス、ニジェールサウルス、クリプトプス、ンクウェバサウルス、パラントドンは、アフリカで知られている前期白亜紀の恐竜です。前期白亜紀は、アフリカが最終的に南アメリカから分離し、南大西洋を形成した時期であったため、アフリカの恐竜にとって重要な時期でした。アフリカの恐竜が孤立したために固有性を発達させ始めたので、これは重要なイベントでした。アフリカの後期白亜紀は主に北アフリカから知られています。白亜紀後期の初期には、北アフリカには恐竜の動物相が豊富に生息していました。スピノサウルス、カルカロドントサウルス、ルゴプス、バハリサウルス、デルタドロメウス、パラリティタン、アエギプトサウルス、オウラノサウルスが含まれます。
コンテンツ
1 含めるための基準
2 アフリカの恐竜のリスト
2.1 有効な属 2.2 疑わしい、不確実な、無効な属
3 タイムライン
4 も参照してください
5 参考文献
含めるための基準
クリーチャーは恐竜属のリストに表示されている必要が
生き物の化石はアフリカで発見されたに違いありません。
このリストは、Category:DinosaursofAfricaを補完するものです。
アフリカの恐竜のリスト編集
有効な属
名前 年 形成
位置
ノート
画像
アールドニクス 2010年
エリオット層(前期ジュラ紀、シネムーリアン)![]()
南アフリカ 主に二足歩行ですが、四足歩行も可能です
アブリクトサウルス 1975
エリオット層(前期ジュラ紀、ヘッタンギアンからシネムーリアン)![]()
南アフリカ 2つの頭蓋骨から知られており、そのうちの1つは牙を持っています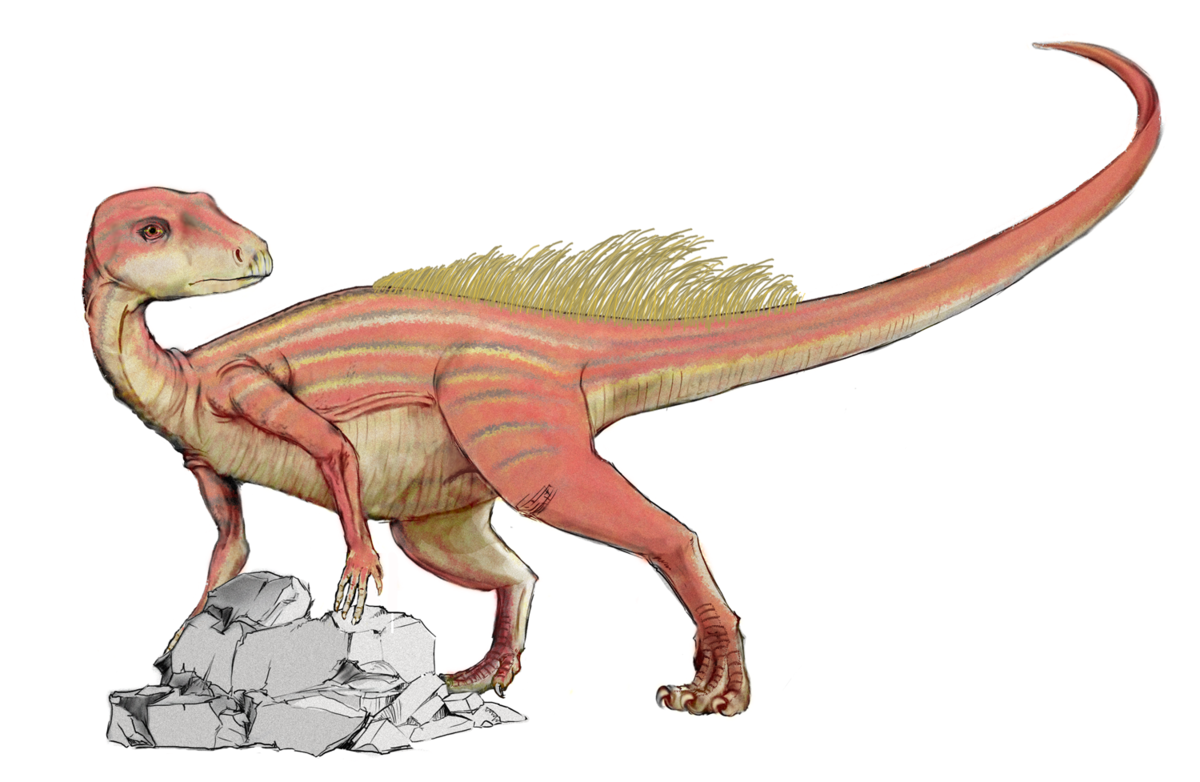
Adratiklit 2019年
エルメルスII層(中期ジュラ紀、バトニアン
![]()
モロッコ 最も古い既知のステゴサウルス。幼い頃にも関わらず、ジュラ紀後期のヨーロッパの形態に関連している
アエギプトサウルス 1932年
バハリヤ層(白亜紀後期、セノマニアン)![]()
エジプト そのホロタイプ標本は第二次世界大戦中に失われました
アフロミムス 2017年
エルハズ層(前期白亜紀、アプチアンからアルビアン)![]()
ニジェール もともとはオルニトミモサウルスと記述されていましたが、後にノアサウルスと再記述されました。
アフロベナトル 1994年
ティユラレン層(中期ジュラ紀、バトニアン)![]()
ニジェール もともとは前期白亜紀から雹を降らせると考えられていました
アジナビア 2020
Ouled Abdoun Basin(白亜紀後期、マーストリヒチアン)![]()
モロッコ アフリカで知られている唯一のハドロサウルス。それ以外の場合はヨーロッパ固有のランベオサウルスの分岐群の一部
アルゴアサウルス 1904年
カークウッド層(前期白亜紀、ベリアシアンからバランギニアン)![]()
南アフリカ 今日、ほんの数個の骨から知られています。研究する前に、さらにいくつかがレンガにされた可能性があります。
アンゴラティタン 2011
イトンベ層(白亜紀後期、コニアシアン)![]()
アンゴラ アンゴラから記述された最初の非鳥類恐竜
アンテトニトルス 2003年
エリオット層(前期ジュラ紀、ヘッタンギアン)![]()
南アフリカ 体重を支えるのに適した前肢と足をつかんでいた
アルクサウルス 2011
エリオット層(前期ジュラ紀、プリンスバッキアン)![]()
南アフリカ 基礎竜脚形亜目と派生竜脚形亜目の両方の所有特性
アトラサウルス 1999年
Guettioua累層(中期ジュラ紀、バトニアンからカロビアン)![]()
モロッコ 特徴的に細長い脚を持つ竜脚類
アウストラロドクス 2007年
テンダグル層(ジュラ紀後期、チトニアン)![]()
タンザニア 潜在的に最も古い既知のeuhelopodid 
バハリサウルス 1934年
バハリヤ層(白亜紀後期、セノマニアン)![]()
エジプト 大きいが、ごく少数の遺跡から知られている
ベルベロサウルス 2007年
アジラル層(前期ジュラ紀、トアルシアン)![]()
モロッコ 中型の初期のケラトサウルス
ブリカナサウルス 1985
エリオット層(三畳紀後期、ノリアン)![]()
南アフリカ ニッチが他の後期三畳紀のエリオット竜脚形亜目と分割された「超堅牢」な形態
カルカロドントサウルス 1931年
バハリヤ層、コンチネンタルインターカレア、エクハー累層、ケムケム累層(白亜紀後期、セノマニアン)![]()
アルジェリア
![]()
エジプト
![]()
モロッコ
![]()
ニジェール ティラノサウルスやギガノトサウルスに匹敵する、最も長い肉食恐竜の1つ。2つの種が知られています
シェブサウルス 2005年
アイッサ層(中期ジュラ紀、カロビアン)![]()
アルジェリア 2つの幼体から知られている
チェナニサウルス 2017年
Ouled Abdoun Basin(白亜紀後期、マーストリヒチアン)![]()
モロッコ 大きくて生き残ったアベリサウルス
クリスタトゥサウルス 1998年
エルハズ層(前期白亜紀、アプチアンからアルビアン)![]()
ニジェール スコミムスの同義語である可能性がありますが、有効な属である可能性もあります。
デルタドロメウス 1996年
ケムケムグループ(白亜紀後期、セノマニアン)![]()
モロッコ 基礎コエルロサウルス、カルノサウルス、メガラプトル類、、そして最近では巨大なノアサウルスであることがさまざまに示唆されています。
ディクラエオサウルス 1914年
テンダグル層(ジュラ紀後期、キンメリッジアンからチトニアン)![]()
タンザニア 低いブラウザであったかもしれない短い首の竜脚類
ドラコヴェナトル 2005年
エリオット層(前期ジュラ紀、ヘッタンギアン)![]()
南アフリカ 頭蓋骨の断片からのみ知られていますが、それらはそれがディロフォサウルスに関連していることを伝えるのに十分です

ジサロトサウルス 1919年
テンダグル層(ジュラ紀後期、キンメリッジアン)![]()
タンザニア その生活史、 食事、そして病気について多くを明らかにした複数の遺跡から知られています。
エラフロサウルス 1920年
テンダグル層(ジュラ紀後期、キンメリッジアン)![]()
タンザニア 大きな基底ケラトサウルス、潜在的にノアサウルス、繊細な体格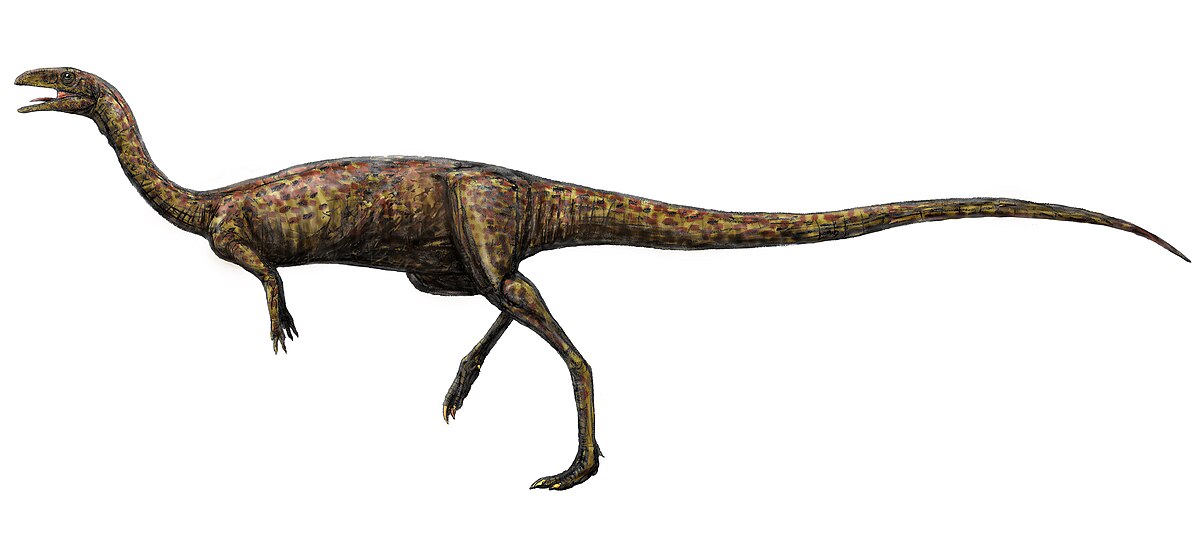
エルハゾサウルス 2009年
エルハズ層(前期白亜紀、アプチアン)![]()
ニジェール ヴァルドサウルスと密接な関係

エオカルカリア 2008年
エルハズ層(前期白亜紀、アルビアン)![]()
ニジェール その前頭骨は太い帯に腫れ、それはそれに威嚇するようなまぶしさを与えました
エオクルソル 2007年
エリオット層(前期ジュラ紀、ヘッタンギアン)![]()
南アフリカ 最も完全に知られている初期の鳥盤類の1人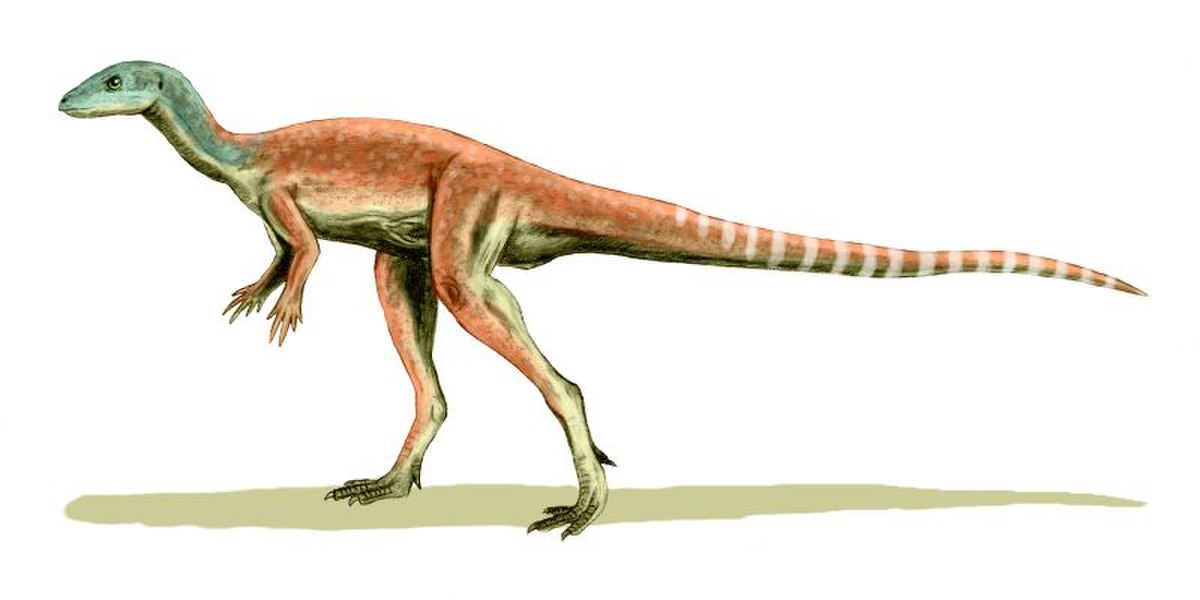
エウクネメサウルス 1920年
エリオット層(三畳紀後期、カーニアン期からノリアン期)![]()
南アフリカ この竜脚形亜目のいくつかの化石は、もともと巨大なヘレラサウルスの化石として解釈されていました
ユースケロサウルス 1866年
エリオット層(三畳紀後期、ノリアンからレーティアン)![]()
レソト
![]()
南アフリカ
![]()
ジンバブエ 大きくて頑丈に作られています
ゲラノサウルス 1911年
クラレンス層(前期ジュラ紀、プリンスバッキアンからトアルシアン)![]()
南アフリカ あまり知られていませんが、ヘテロドントサウルスの可能性があります
ギラッファティタン 1988年
テンダグル層(ジュラ紀後期、キンメリッジアンからチトニアン)![]()
タンザニア いくつかの標本から知られています。一般的にブラキオサウルスに関連していますが、2つの間にいくつかの違いが存在します。
グリポニックス 1911年
エリオット層(前期ジュラ紀、ヘッタンギアンからシネムーリアン)![]()
南アフリカ Massospondylusの同義語である可能性がありますが、遠い関係にある可能性もあります。
ヘテロドントサウルス 1962年
クラレンス層、エリオット層(前期ジュラ紀、ヘッタンギアンからプリンスバッキアン)![]()
南アフリカ 切歯や牙の類似物、角質のくちばしなど、3種類の歯を持っています
イグナヴサウルス 2010年
エリオット層(前期ジュラ紀、ヘッタンギアン)![]()
レソト Massospondylus の幼体と考えられることもありますが、サラサウルスとより密接に関連している可能性が
イノサウルス 1960年
Irhazer Group?(前期白亜紀、ベリアシアンからバレミアン)?![]()
ニジェール あまり知られていない
ジャネンシア 1991
テンダグル層(ジュラ紀後期、キンメリッジアンからチトニアン)![]()
タンザニア 時折ティタノサウルスと考えられますが、いくつかの分析では、ベルサウルスとハエスタサウルス のあるクレードで、トゥリアサウリアとの親和性があり、より基本的な真竜脚類として回復しています。
ジョバリア 1999年
ティユラレン層(中期ジュラ紀、バトニアン)![]()
ニジェール ほぼ完全なスケルトンから知られています
カングナサウルス 1915年
カラハリ鉱床形成(前期白亜紀、アプチアン)![]()
南アフリカ ドリオサウルスとの比較が行われていますが、ある研究ではエラスマリア内での位置が示唆されています。
カロンガサウルス 2005年
恐竜のベッド(前期白亜紀、アプチアン)![]()
マラウイ 下顎と歯だけから説明
ケントロサウルス 1915年
テンダグル層(ジュラ紀後期、キンメリッジアンからチトニアン)![]()
タンザニア 尾に向かって徐々にスパイクに融合する2列のプレートと、各肩に長いスパイクを所有していました
コルモルモ 2020
エリオット層(三畳紀後期、ノリアン)![]()
レソト 正式な説明の前は、非公式に「Kholumolumosaurus」および「Thotobolosaurus」と呼ばれていました。
クリプトプス 2008年
エルハズ層(前期白亜紀、アプチアンからアルビアン)![]()
ニジェール このアベリサウルスと呼ばれる頭蓋後の遺体は、代わりにカルカロドントサウルスに由来する可能性があります。
レドマハディ 2018年
エリオット層(前期ジュラ紀、ヘッタンギアンからシネムーリアン)![]()
南アフリカ 重量が12トン(26,000ポンド)と推定される最大の三畳紀恐竜の1つ
レソトサウルス 1978年
エリオット層(前期ジュラ紀、ヘッタンギアンからプリンスバッキアン)![]()
レソト
![]()
南アフリカ 複数の個人の遺骨が発見されており、群生するライフスタイルを示唆しています
ルルドゥサウルス 1999年
エルハズ層(前期白亜紀、アルビアン)![]()
ニジェール その体と手足の比率は、それがカバに似た半水生草食動物であったことを示唆しています。
リコリヌス 1924年
エリオット層(前期ジュラ紀、ヘッタンギアンからシネムーリアン)![]()
南アフリカ もともとキノドン類と誤認されていた
マラウィサウルス 1993年
恐竜のベッド(前期白亜紀、バレミアンからアプチアン)![]()
マラウイ 頭蓋骨と皮骨の材料が知られています
マンスウラサウルス 2018年
Quseir層(白亜紀後期、カンパニアン)![]()
エジプト ユーラシアの形態と密接に関連しています。その発見は、アフリカが白亜紀後期に以前考えられていたほど孤立していなかったことを示している
Massospondylus 1854年
ブッシュフェルド砂岩、クラレンス層、エリオット層、森林砂岩(前期ジュラ紀、ヘッタンギアンからプリンスバッキアン)![]()
レソト
![]()
南アフリカ
![]()
ジンバブエ 豊富な遺跡が発見されました。いくつかの標本はかつて彼ら自身の属と種に割り当てられました
メラノロサウルス 1924年
エリオット層(三畳紀後期、ノリアン)![]()
南アフリカ 丈夫な四足動物の草食動物。この属に割り当てられたいくつかの標本は、同じ分類群を表していない可能性があります
メロクテノス 2016年
エリオット層(三畳紀後期、ノリアンからレーティアン)![]()
レソト その大腿骨は異常に頑丈でした
Mnyamawamtuka 2019年
ガルラ層(前期白亜紀、アプチアン)![]()
タンザニア moyowamkiaの具体的な名前は「ハートテール」のスワヒリ語で、尾椎のハート型の断面を指します。
ングウェヴ 2019年
エリオット層(前期ジュラ紀、ヘッタンギアンからシネムーリアン)![]()
南アフリカ もともとマッソスポンディルスに割り当てられた頭蓋骨から知られており、その独特の比率に基づいて独自の属に割り当てられています
ニジェールサウルス 1999年
エルハズ層(前期白亜紀、アプチアンからアルビアン)![]()
ニジェール その歯はすべて顎の前にあり、頭蓋骨の他の部分よりも幅が広く、低ブラウジングに適応していました。
ンクウェバサウルス 2000年
カークウッド層(前期白亜紀、ベリアシアンからバランギニアン)![]()
南アフリカ アフリカ本土にちなんで名付けられた最初のコエルロサウルス
オロサウルス 1867年
エリオット層(三畳紀後期、カーニアン期からノリアン期)![]()
南アフリカ おそらくEuskelosaurusの同義語
オスタフリカサウルス 2012年
テンダグル層(ジュラ紀後期、チトニアン)![]()
タンザニア 単一の歯から初期スピノサウルスとして説明されていますが、ケラトサウルスである可能性も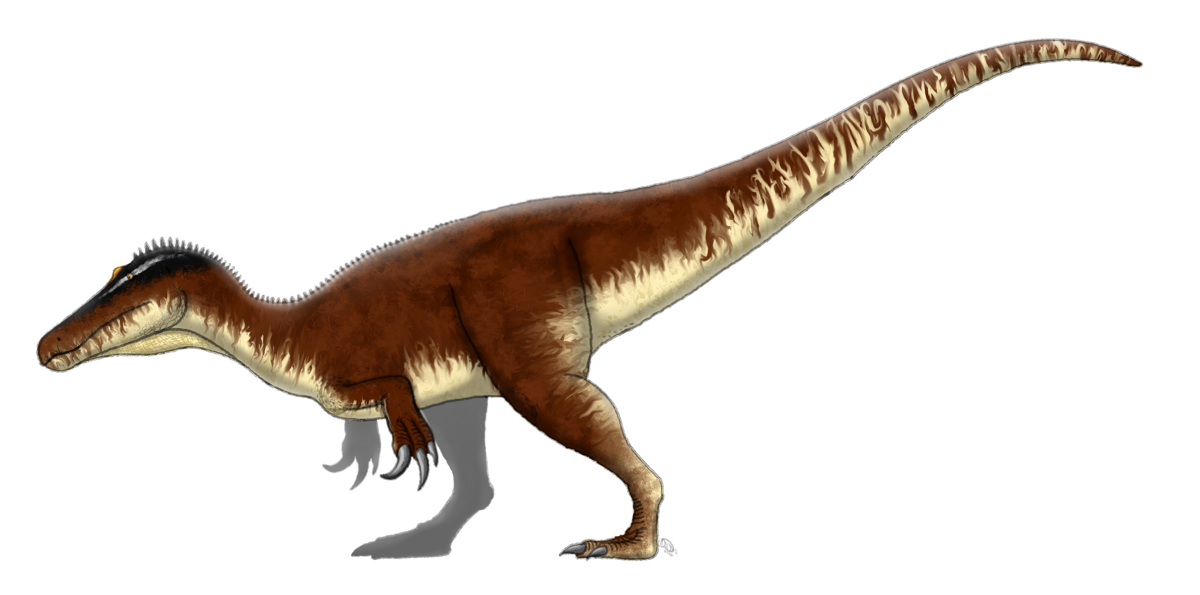
オウラノサウルス 1976年
エルハズ層(前期白亜紀、アプチアンからアルビアン)![]()
ニジェール 椎骨から突き出た長い神経棘は、帆やこぶを支えていた可能性があります
パラリティタン 2001年
バハリヤ層(白亜紀後期、セノマニアン)![]()
エジプト マングローブの沼に住んでいただろう
パラントドン 1929年
カークウッド層(前期白亜紀、ベリアシアンからバランギニアン)![]()
南アフリカ 断片的な標本からしか知られていませんが、それがステゴサウルスであったことを伝えるのに十分です
ペゴマスタックス 2012年
エリオット層(前期ジュラ紀、ヘッタンギアンからシネムーリアン)![]()
南アフリカ 短いくちばしで頑丈な顎を持っていた
Plateosauravus 1932年
エリオット層(三畳紀後期、ノリアン)![]()
南アフリカ 少年の標本を含む複数の標本から知られています
プラネサウラ 2015年
エリオット層(前期ジュラ紀、ヘッタンギアンからシネムーリアン)![]()
南アフリカ 四足歩行の短い首の低いブラウザ
レッバキサウルス 1950年
アウフー層(白亜紀後期、セノマニアン)![]()
モロッコ 背の高い尾根や低い帆を支えていたであろう細長い神経棘の列を運んだ
ルゴプス 2004年
エクハー累層(白亜紀後期、セノマニアン)![]()
ニジェール 頭蓋骨の上部にある2列の7つの穴を保持します。これは、ディスプレイ構造を固定している可能性があります。
ルクワティタン 2014年
ガルラ層(前期白亜紀、アルビアン)![]()
タンザニア 白亜紀中期のアフリカで知られている数少ないティタノサウルスの1つで、進化の歴史のギャップを埋めています
セファパノサウルス 2015年
エリオット層(前期ジュラ紀、ヘッタンギアン)![]()
南アフリカ 独特の十字型のレンゲがありました
シンゴパナ 2017年
ガルラ層(前期白亜紀、アルビアン)![]()
タンザニア 南米のティタノサウルスと最も密接に関連している
スピコメルス 2021年
エルマースグループ(中期ジュラ紀、バトニアンからカロビアン)![]()
モロッコ 知られている最古の曲竜類であり、アフリカからの最初のもの。独特なことに、その皮骨はその肋骨に直接融合していました
スピノフォロサウルス 2009年
Irhazer Shale(中期ジュラ紀、バトニアン)![]()
ニジェール もともとは剣竜類のようなサゴマイザーを所有していると説明されていました。これらは鎖骨であることが判明した。肩が高く首が高い高ブラウザ
スピノサウルス 1915年
バハリヤ層、シュニーニ層、ケムケムグループ(白亜紀後期、セノマニアンからチューロニアン)![]()
エジプト
![]()
モロッコ
![]()
チュニジア 水かきのある足やパドルのような尾など、半水生のライフスタイルを示唆する無数の機能を備えています。
スピノストロフェウス 2004年
ティユラレン層(中期ジュラ紀、バトニアン)![]()
ニジェール おそらくエラフロサウルスと密接に関連している
スコミムス 1998年
エルハズ層(前期白亜紀、アプチアンからアルビアン)![]()
ニジェール バリオニクスに似ていますが、背中に低い帆があります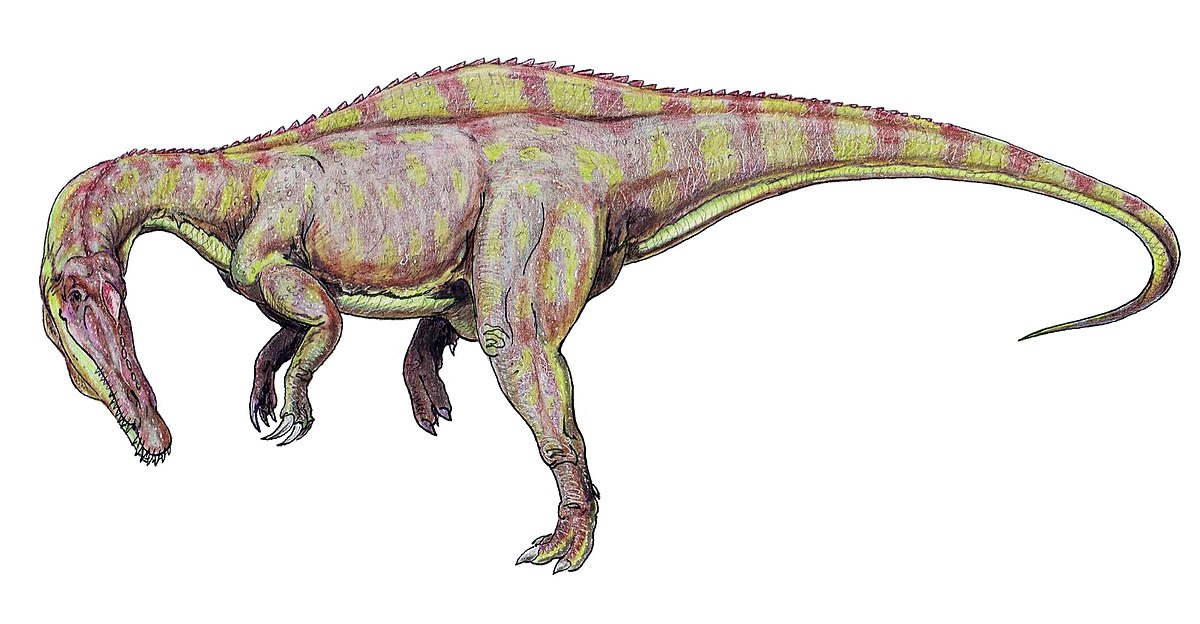
タタウイネア 2013年
AïnelGuettar累層(前期白亜紀、アルビアン)![]()
チュニジア その骨は広範囲に空気圧がかけられており、竜脚類は鳥のような呼吸器系を持っているという理論を支持しています
タゾウダサウルス 2004年
アジラル層(前期ジュラ紀、トアルシアン)![]()
モロッコ 適度に完全な遺跡から知られている数少ない前期ジュラ紀の竜脚類の1つ
テンダグリア 2000年
テンダグル層(ジュラ紀後期、チトニアン)![]()
タンザニア トゥリアサウリアとの親和性がありそうな大型竜脚類
トルニエリア 1911年
テンダグル層(ジュラ紀後期、チトニアン)![]()
タンザニア その歴史を通して異なる属に割り当てられてきました
ヴェテルプリスティサウルス 2011
テンダグル層(ジュラ紀後期、キンメリッジアンからチトニアン)![]()
タンザニア アクロカントサウルスの大規模な初期の親戚

ヴルカノドン 1972年
森林砂岩(前期ジュラ紀、シネムーリアンからプリンスバッキアン)![]()
ジンバブエ 獣脚類の歯はホロタイプに関連していることがわかった
Wamweracaudia 2019年
テンダグル層(ジュラ紀後期、チトニアン)![]()
タンザニア アジア以外から知られている最初の決定的なマメンチサウルス
疑わしい、不確実な、無効な属
Aetonyx palustris : Massospondylusの潜在的なジュニアシノニム。
Fabrosaurus australis:ファブロサウルスと呼ばれる小さな鳥盤類の家族の一部と見なされることもレソトサウルスの同義語である可能性が
Gigantoscelus molengraafi:おそらくEuskelosaurusの同義語ですが、確認できません。
ジポサウルス:現代の竜脚形亜目の幼体である可能性が中国の種も命名されていますが、この属に属していない可能性が
Hortalotarsus skirtopodus: Massospondylusの同義語の可能性。
「Likhoelesaurus」:巨大な初期のカルノサウルスであることが示唆されていますが、実際には偽鰐類である可能性が
メガプノサウルス:アフリカの種M. rhodesiensisは、コエロフィシス属に属する可能性が別の参照種であるM.kayentakataeには、おそらく独自の属名が必要です。
Nyasasaurus parringtoni:2013年に、アニシアンにまでさかのぼる最も古い既知の恐竜として説明されていますが、この日付とその分類の両方が疑問視されています。
Sauroniops pachytholus :単一の肥厚した前頭葉からのみ知られている、カルカロドントサウルスと同義である可能性が
Sigilmassaurus brevicollis:スピノサウルスと同時代のスピノサウルス; 別個の属および/または種としてのその状態は不確かです。
トルボサウルス:元々メガロサウルスインゲンと名付けられた歯は、この属に再割り当てされました。
タイムライン
これは、上記のリストから選択した恐竜のタイムラインです。時間はx軸に沿ってmyaで測定されます。
も参照してください
アフリカの鳥のリスト
参考文献
![]()
恐竜ポータル
![]()
アフリカポータル
^ Maidment、Susannah CR; レイヴン、トーマスJ .; Ouarhache、Driss; バレット、ポールM.(2019-08-16)。「北アフリカ初の剣竜類:ゴンドワナ大陸の装盾亜目恐竜の多様性への影響」。ゴンドワナリサーチ。77:82–97。土井:10.1016/j.gr.2019.07.007。ISSN1342-937X 。_ ^ Sereno、P.(2017)。「アフリカからの前期白亜紀のオルニトミモサウルス(恐竜:コエルロサウルス)」。Ameghiniana。54(5):576–616。土井:10.5710/AMGH.23.10.2017.3155。S2CID134718338。_
^ セローニ、マサチューセッツ州; フロリダ州アグノリン; BrissónEgli、F .; Novas、FE(2019)。「AfromimustenerensisSerenoの系統発生的位置、2017年およびその古生物地理学的意味」。アフリカ地球科学ジャーナル。159:103572。doi : 10.1016 /j.jafrearsci.2019.103572。
^ Longrich、Nicholas R .; Suberbiola、Xabier Pereda; パイロン、R。アレクサンダー; Jalil、Nour-Eddine(2020)。「アフリカからの最初のアヒルの子恐竜(ハドロサウルス科:ランベオサウルス科)と恐竜の生物地理学における海洋分散の役割」。白亜紀の研究:104678。doi : 10.1016 /j.cretres.2020.104678。
^ ドン・レッセム; ドナルドF.グラット(1993)。恐竜協会の恐竜百科事典。トレーシーフォード(イラスト)…; 科学顧問、ピーター・ドッドソン(第1版)。ニューヨーク:ランダムハウス。p。 16。ISBN 0-679-41770-2。
^ マニオン、PD; Upchurch、P .; Schwarz、D .; ウィングス、O。(2019)。「タンザニアの後期ジュラ紀テンダグル層からの推定上のチタノサウルスの分類学的親和性:真竜脚類恐竜の進化に対する系統発生的および生物地理学的意味」。リンネ協会の動物学ジャーナル。185(3):784–909。土井:10.1093 / zoolinnean/zly068。hdl:10044/1/64080。
^ Mcphee、Blair W .; Bordy、Emese M .; シシオ、ララ; Choiniere、Jonah N.(2017)。「アフリカ南部のエリオット層の竜脚形亜目生層序:三畳紀-ジュラ紀境界を越えた竜脚形亜目の進化の追跡」。ActaPalaeontologicaPolonica。62(3):441–465。土井:10.4202/app.00377.2017。
^ レーセルダ、マウロBS; グリロ、オーランドN .; ロマーノ、ペドロSR(2021)。「スピノサウルス科(獣脚類、メガロサウルス科)の吻側形態:前上顎骨の形状変化と新しい系統学的推論」。歴史生物学:1–21。土井:10.1080/08912963.2021.2000974。S2CID244418803。_ ^ Sereno Dutheil; Iarochene Larsson; リヨンマグウェン; Sidor Varricchio; ウィルソン(1996)。「サハラと白亜紀後期の動物相の分化からの略奪的な恐竜」(PDF)。科学。272(5264):986–991。Bibcode:1996Sci…272..986S。土井:10.1126/science.272.5264.986。PMID8662584。_ S2CID39658297。_
^ SebastiánApesteguía; ネイサンD.スミス; ルベン・フアレス・バリエリ; ピーター・J・マコヴィッキー(2016)。「アルゼンチン、パタゴニアの白亜紀後期からのジダクチルマヌスを持つ珍しい新しい獣脚類」。PLOSONE。11(7):e0157793。Bibcode:2016PLoSO..1157793A。土井:10.1371/journal.pone.0157793。PMC4943716。_ PMID27410683。_ ^ マティアスJ.モッタ; Alexis M. Aranciaga Rolando; SebastiánRozadilla; フェデリコE.アグノリン; ニコラスR.チメント; フェデリコブリッソンエグリ&フェルナンドE.ノバス(2016)。「アルゼンチン、パタゴニア北西部の白亜紀後期(Huincul累層)からの新しい獣脚類動物相」。ニューメキシコ自然史科学博物館紀要。71:231–253。
^ イブラヒム、ニザル; セレノ、ポールC .; Varricchio、David J .; マーティル、デビッドM .; Dutheil、Didier B .; Unwin、David M .; Baidder、Lahssen; ラーソン、ハンスCE; Zouhri、Samir; Kaoukaya、Abdelhadi(2020-04-21)。「モロッコ東部の白亜紀後期のケムケムグループの地質学と古生物学」。ZooKeys(928):1–216。土井:10.3897/zookeys.928.47517。ISSN1313-2970。_ PMC7188693。_ PMID32362741。_ ^ Hübner、TR(2012)。Laudet、V.(ed。)「Dysalotosauruslettowvorbecki(鳥盤類:イグアノドン類)の骨組織学-変動、成長、および影響」。PLOSONE。7(1):e29958。Bibcode:2012PLoSO…729958H。土井:10.1371/journal.pone.0029958。PMC3253128。_ PMID22238683。_ ^ Hübner、TR; Rauhut、OWM(2010)。「Dysalotosauruslettowvorbecki(鳥盤類:イグアノドン類)の幼い頭蓋骨、および鳥脚類恐竜における頭蓋個体発生、系統発生、および分類法への影響」。リンネ協会の動物学ジャーナル。160(2):366–396。土井:10.1111/j.1096-3642.2010.00620.x。
^ Witzmann、F .; クレソン、KM; ハンペ、O .; Wieder、F .; ヒルガー、A .; マンケ、私。ニーダーハーゲン、M .; ロスチャイルド、BM; Asbach、P.(2011)。「ジュラ紀の恐竜の骨のパジェット病」。カレントバイオロジー。21(17):R647–8。土井:10.1016/j.cub.2011.08.006。PMID21920291。_ ^ Rauhut、OWM、およびCarrano、MT(2016)。タンザニアのテンダグルの後期ジュラ紀からの獣脚類恐竜エラフロサウルスバンベルギジャネンシュ、1920年。リンネ協会の動物学ジャーナル、(事前オンライン出版) doi: 10.1111 / zoj.12425 ^ テイラー、MP(2009)。「 BrachiosaurusaltithoraxRiggs1903 (恐竜、竜脚類)の再評価と、 Giraffatitan brancai(Janensch 1914)からの一般的な分離」 (PDF)。Journal ofVertebratePaleontology。29(3):787–806。土井:10.1671/039.029.0309。S2CID15220647。_
^ イェーツ、AM; ボンナン、MF; Neveling、J .; Chinsamy、A .; 黒ひげ、MG(2010)。「南アフリカの前期ジュラ紀からの新しい過渡的な竜脚形恐竜と竜脚類の摂食と四足動物の進化」。王立協会紀要B。277(1682):787–794。土井:10.1098/rspb.2009.1440。PMC2842739。_ PMID19906674。_ ^ イェーツ、アダムM .; マシューF.ボナン; Johann Neveling(2011)。「南アフリカの前期ジュラ紀からの新しい基本的な竜脚形恐竜」。Journal ofVertebratePaleontology。31(3):610–625。土井:10.1080/02724634.2011.560626。S2CID85589914。_ ^ Chapelle KEJ、Barrett PM、Botha J、Choiniere JN 2019. Ngwevu intloko:南アフリカの下部ジュラ紀エリオット層からの新しい初期竜脚形恐竜とMassospondyluscarinatusの頭蓋個体発生についてのコメント。PeerJ 7:e7240 https://doi.org/10.7717/peerj.7240 ^ Wild、R.(1991)。Janenschia ng robusta(E. Fraas 1908)pro Tornieria robusta(E. Fraas 1908)(Reptilia、Saurischia、Sauropodomorpha)[Janenschia ng robusta(E. Fraas 1908)for Tornieria robusta(E. Fraas 1908)(Reptilia、Saurischia、Sauropodomorpha )]。StuttgarterBeiträgezurNaturkunde、Serie B(GeologieundPaläontologie)173:1–4 ^ ダニエラシュワルツ; フィリップマニオン; オリバーウィングス; クリスチャンマイヤー(2020)。「竜脚類恐竜アマンジア(「オルニトプシス/ケティオサウリスクス」)greppini n。gen 。およびその他の脊椎動物の再記述は、スイス、ムーティエのキンメリッジアン(後期ジュラ紀)ロイシェネット層から残っています。地球科学のスイスジャーナル。土井:10.1186/s00015-020-00355-5。
^ Ruiz-Omeñaca、JoséIgnacio; ペレダ・スベルビオラ、ザビエル; ガルトン、ピーターM.(2007)。「カロヴォサウルスleedsi、イギリスの中期ジュラ紀からの最も初期のドリオサウルス恐竜(鳥盤類:Euornithopoda)」。カーペンターケネス(編)。角とくちばし:角竜類と鳥脚類の恐竜。ブルーミントンとインディアナポリス:インディアナ大学出版局。pp。3–16。ISBN 978-0-253-34817-3。
^ デュドネ、P.-E .; Cruzado-Caballero、P .; ゴドフロイト、P .; トルトサ、T。(2020-07-20)。「角脚類恐竜の新しい系統発生」(PDF)。歴史生物学。33(10):2335–2355。土井:10.1080/08912963.2020.1793979。ISSN0891-2963。_ S2CID221854017。_
^ Carrano、Matthew T .; ロジャーBJベンソン; スコットD.サンプソン(2012)。「テタヌラ類(恐竜:獣脚類)の系統発生」。系統的古生物学ジャーナル。10(2):211–300。土井:10.1080/14772019.2011.630927。
^ McPhee、Blair W .; ベンソン、ロジャーBJ; Botha-Brink、Jennifer; Bordy、Emese M.&Choiniere、Jonah N.(2018)。「南アフリカの初期のジュラ紀からの巨大な恐竜と初期の竜脚形亜目における四肢への移行」。カレントバイオロジー。28(19):3143–3151.e7。土井:10.1016/j.cub.2018.07.063。PMID30270189。_ ^ TR、ホルツ、ジュニア; レイ、L。(2007)。恐竜:すべての年齢の恐竜愛好家のための最も完全な最新の百科事典。ニューヨーク:ランダムハウス。p。257. ISBN 9780375824197。
^ Sallam、H .; Gorscak、E .; オコナー、P .; El-Dawoudi、I .; El-Sayed、S .; セイバー、S。(2017-06-26)。「新しいエジプトの竜脚類は、ヨーロッパとアフリカの間の白亜紀後期の恐竜の分散を明らかにします」。自然。2(3):445–451。土井:10.1038/s41559-017-0455-5。PMID29379183。_ ^ ビュフェトー、エリック(2012)。「テンダグル(タンザニア)の後期ジュラ紀からの初期スピノサウルス恐竜とスピノサウルス歯列の進化」(PDF)。Oryctos。10:1〜8。
^ ソト、マティアス; トリニョ、パブロ; ペレア、ダニエル(2020-11-01)。「タクアレンボ層(後期ジュラ紀、ウルグアイ)からのケラトサウルス(獣脚類、ケラトサウルス)の歯」。南アメリカの地球科学のジャーナル。103:102781。Bibcode:2020JSAES.10302781S。土井:10.1016/j.jsames.2020.102781。ISSN0895-9811。_ S2CID224842133。_ ^ セレノ、ポールC .; ウィルソン、ジェフリーA .; コンラッド、ジャックL.(2004-07-07)。「新しい恐竜は白亜紀中期の南部の陸塊をつなぐ」。王立協会の議事録B:生物科学。271(1546):1325–1330。土井:10.1098/rspb.2004.2692。ISSN0962-8452。_ PMC1691741。_ PMID15306329。_ ^ Remes、K .; Ortega、F .; フィエロ、I .; Joger、U .; コスマ、R .; マリン・フェラー、JM; 井出、OAu; マガ、A .; ファルケ、AA(2009)。「ニジェールの中期ジュラ紀からの新しい基本的な竜脚類恐竜と竜脚類の初期の進化」。PLOSONE。4(9):e6924。Bibcode:2009PLoSO…4.6924R。土井:10.1371/journal.pone.0006924。PMC2737122。_ PMID19756139。_ ^ Tschopp、E .; マテウス、O。(2013)。「竜脚類恐竜の鎖骨、鎖骨間、腹肋骨、および胸骨肋骨:ディプロドクス科からの新しい報告とそれらの形態学的、機能的および進化的意味」。解剖学ジャーナル。222(3):321–340。土井:10.1111/joa.12012。PMC3582252。_ PMID23190365。_ ^ Vidal、D .; モチョ、P .; Aberasturi、A .; Sanz、JL; Ortega、F.(2020)。「スピノフォロサウルスの高度なブラウジング骨格の適応は、竜脚類恐竜の進化的革新を明らかにします」。ScientificReports。10(1):6638。Bibcode:2020NatSR..10.6638V。土井:10.1038/s41598-020-63439-0。PMC7171156。_ PMID32313018。_ S2CID215819745。_ ^ イブラヒム、ニザル; セレノ、ポールC .; ダルサッソ、クリスティアーノ; マガヌコ、シモーネ; ファブリ、マッテオ; マーティル、デビッドM .; Zouhri、Samir; ネイサン・ミルボルド; Lurino、Dawid A.(2014)。「巨大な略奪恐竜における半水生の適応」。科学。345(6204):1613–6。Bibcode:2014Sci…345.1613I。土井:10.1126/science.1258750。PMID25213375。_ S2CID34421257。_ 補足情報 ^ イブラヒム、ニザル; マガヌコ、シモーネ; ダルサッソ、クリスティアーノ; Fabbri、Matteo; オーディトーレ、マルコ; Bindellini、Gabriele; マーティル、デビッドM .; Zouhri、Samir; Mattarelli、Diego A .; Unwin、David M .; ウィーマン、ジャスミナ(2020)。「獣脚類恐竜の尾推進水泳運動」。自然。581(7806):67–70。Bibcode:2020Natur.581…67I。土井:10.1038/s41586-020-2190-3。ISSN1476-4687。_ PMID32376955。_ S2CID216650535。_ ^ ディエゴ・ポル&オリバーWMラウフト(2012)。「パタゴニアの中期ジュラ紀のアベリサウルスと獣脚類恐竜の初期の多様化」。王立協会の議事録B:生物科学。279(1804):3170–5。土井:10.1098/rspb.2012.0660。PMC3385738。_ PMID22628475。_ “