Mearns_coyote#Subspecies
その他の用法については「コヨーテ 」をご覧
コヨーテ( Canis latrans ) は、北米原産の犬種です。近縁種のオオカミよりも小さく、近縁種の東オオカミやアカオオカミよりもわずかに小さい。イヌジャッカルがユーラシアで行うのと同じ生態学的ニッチの多くを満たしています。コヨーテはより大きく、より捕食性が高く、かつて行動生態学者によってアメリカのジャッカルと呼ばれていました。種の他の歴史的な名前には、プレーリー ウルフとブラッシュ ウルフが含まれます。
コヨーテ
時間範囲:中期更新世– 現在 (0.74–0.85 Ma )
マウンテン コヨーテ ( C. l. lestes )
保存状況
軽度懸念 ( IUCN 3.1 )
科学的分類
王国:
動物界
門:
脊索動物
クラス: 哺乳類 注文: 食肉目 家族: イヌ科
属: カニス 種族:
C.ラトランス
二項名 Canis latrans セイ、1823
Canis latransのモダンな範囲 リスト
Canis andersoni Merriam , 1910
カニス・カネロエンシス・スキナー、1942年
Canis clepticusエリオット、1903
Canis estor Merriam , 1897
Canis frustror ウッドハウス、1851
Canis goldmani Merriam , 1904
Canis hondurensis ゴールドマン, 1936
Canis impavidus アレン、1903
Canis irvingtonensis Savage、1951年
Canis jamesi Townsend、1912年
Canis lestes Merriam、1897年
Canis mearnsi Merriam , 1897
Canis microdon Merriam , 1897
Canis nebrascensis Merriam , 1898
Canis ochropus Eschscholtz、1829年
Canis orcutti Merriam , 1910
Canis pallidus Merriam , 1897
Canis peninsulae Merriam , 1897
Canis riviveronis 干し草、1917
Canis vigilis Merriam , 1897
Lyciscus cagottisハミルトン-スミス、1839
コヨーテは、北米全体に広く分布し、豊富であるため、国際自然保護連合によって最も懸念されていないものとしてリストされています。この種は用途が広く、人間によって変更された環境に適応して拡大することができます。米国東部とカナダの都市部に移動することで、その範囲を拡大しています。コヨーテは、2013 年に初めてパナマ東部 (生息域からパナマ運河の向こう側) で目撃されました。
コヨーテには 19 の認識された亜種が平均的な男性の体重は 8 ~ 20 kg (18 ~ 44 ポンド)、平均的な女性の体重は 7 ~ 18 kg (15 ~ 40 ポンド) です。彼らの毛皮の色は、地理によって多少異なりますが、主に明るい灰色と赤、または黒と白が散在するフルブスです。それは社会組織において非常に柔軟であり、家族単位で、または無関係な個人のゆるい編成のパックで生活しています。主に肉食性で、主にシカ、ウサギ、ノウサギ、げっ歯類、鳥類、爬虫類、両生類、魚類、無脊椎動物を主食としていますが、果物や野菜を食べることも特徴的な発声は、孤独な個体による咆哮です。人間はコヨーテの最大の脅威であり、次にクーガーとハイイロオオカミが続きます。それにもかかわらず、コヨーテは灰色オオカミ、東部オオカミ、またはアカオオカミと交尾することがあり、「コイウルフ」ハイブリッドを生み出します. 北アメリカの北東部地域では、イースタン コヨーテ(より大きな亜種ですが、オオカミよりはまだ小さい) は、さまざまな種類のオオカミとのさまざまな歴史的および最近の交配の結果です。遺伝子研究によると、ほとんどの北アメリカのオオカミには、ある程度のコヨーテ DNA が含まれています。
コヨーテは、主にアリドアメリカのネイティブ アメリカンの民間伝承の著名なキャラクターであり、通常、実際のコヨーテまたは人間の形を交互にとるトリックスターとして描かれています。他のトリックスター フィギュアと同様に、コヨーテは欺瞞とユーモアを使って社会的慣習に反抗します。この動物は、メソアメリカの宇宙論では軍事力の象徴として特に尊敬されていました。ヨーロッパによるアメリカ大陸の植民地化の後、英米文化では臆病で信頼できない動物と見なされました。公のイメージが向上したオオカミとは異なり、コヨーテに対する態度は依然として否定的です。
コンテンツ
1 説明
2 分類と進化
2.1 歴史 2.2 命名と語源 2.3 進化
2.3.1 化石記録
2.3.2 DNA証拠
2.4 亜種 2.5 ハイブリダイゼーション
3 行動
3.1 社会的および生殖行動 3.2 縄張り意識と保護行動 3.3 狩猟と摂食行動 3.4 コミュニケーション
3.4.1 ボディランゲージ
3.4.2 発声
4 エコロジー
4.1 生息地 4.2 ダイエット 4.3 敵と競争相手
5 範囲
6 病気と寄生虫
7 人間関係
7.1 民話や神話では 7.2 人間への攻撃 7.3 家畜とペットの捕食 7.4 狩猟 7.5 毛皮の用途 7.6 飼いならしやすさ 7.7 有名メディアで
8 ノート
9 引用
10 一般的な情報源
11 参考文献
11.1 書籍 11.2 ビデオ 11.3 オーディオブック
12 外部リンク
説明

マウンテン コヨーテ ( C. l. lestes ) の頭
のクローズ アップ
コヨーテのオスの平均体重は 8 ~ 20 kg (18 ~ 44 ポンド)、メスの平均体重は 7 ~ 18 kg (15 ~ 40 ポンド) ですが、サイズは地理的に異なります。平均 18 kg (40 ポンド) の北部亜種は、平均 11.5 kg (25 ポンド) のメキシコ南部亜種よりも大きくなる傾向が全長は平均して 1.0 ~ 1.35 m (3 フィート 3 ~ 4 フィート 5 インチ) です。尾の長さは 40 cm (16 インチ) で、メスは体長と身長の両方が短くなっています。記録上最大のコヨーテは、1937 年11 月19 日にワイオミング州アフトンの近くで殺されたオスで 、鼻から尻尾まで 1.5 m (4 フィート 11 インチ)、体重は 34 kg (75 ポンド) でした。臭腺は尾の付け根の上部にあり、青みがかった黒色です。
コヨーテの毛皮の色と質感は、地理的に多少異なります。髪の主な色は明るい灰色と赤、またはフルーブで、体の周りに黒と白が散在しています。高地に生息するコヨーテは、砂漠に生息するコヨーテよりも黒と灰色の色合いが多く、毛羽立ちや白っぽい灰色です。コヨーテの毛皮は、短くて柔らかい下毛と長くて粗い保護毛で構成されています。北部の亜種の毛皮は、南部の亜種よりも長くて密度が高く、一部のメキシコおよび中央アメリカの形態の毛皮は、ほとんど毛むくじゃら (剛毛) です。一般に、大人のコヨーテ (コイウルフ ハイブリッドを含む) は、クロテン色の毛色、濃い新生児の毛色、活動的な上尾腺を備えたふさふさした尾、および白い顔のマスクを持っています。 コヨーテでは白皮症は非常にまれです。 1938 年 3 月 22 日から 1945 年 6 月 30 日までの間に、連邦および協同組合のハンターによって殺された合計 750,000 匹のコヨーテのうち、2 匹だけがアルビノでした。
コヨーテは通常、ハイイロオオカミよりも小さいが、耳が長く、脳蓋が比較的大きく、フレーム、顔、銃口が薄い. 臭腺はオオカミよりも小さいが、色は同じ。毛皮の色のバリエーションはオオカミよりもはるかに少ない。コヨーテはまた、オオカミのように水平ではなく、走ったり歩いたりするときに尻尾を下に運ぶ.
コヨーテの足跡は、犬の足跡とは異なり、より細長く丸みの少ない形をしています。 イヌとは異なり、コヨーテの上部犬歯はオトガイ孔を越えて伸びている。
分類と進化編集
歴史

コヨーテのトルテック絵文字
ヨーロッパ人がアメリカ大陸を植民地化した当時、コヨーテは大部分が大陸の西半分の開けた平原と乾燥地帯に限定されていました。コロンブス以降の初期の歴史的記録では、筆者がコヨーテとオオカミのどちらを描写しているかを判断することはしばしば困難です。イリノイ州カスカスキアで1750 年に地元の司祭によって書かれたある記録によると、そこで遭遇した「オオカミ」はヨーロッパのオオカミよりも小さく、大胆さが劣っていました。エドワーズ郡の 1800 年代初頭の別の報告では、夜に吠えるオオカミについて言及されていましたが、これらはおそらくコヨーテでした。この種は、ルイスとクラークの遠征(1804–1806)の間に数回遭遇したが、ミズーリ川上流のヨーロッパの貿易業者にはすでによく知られていた. 1805 年 5 月 5 日にモンタナ州北東部で執筆したメリウェザー ルイスは、コヨーテについて次のように説明しています。
大草原の小さなオオカミや穴掘り犬は、ほとんど常に開けた平野に住んでいます。彼らは通常、10または12のバンドで、時にはそれ以上のグループで関連付けられ、ゲームが頻繁に行われるパスまたは場所の近くに穴を開けます。単独でシカやヤギを捕まえることができないため、単独で見つかることはめったになく、集団で狩りをします。彼らは巣穴の近くで獲物を頻繁に見て捕まえます。これらの巣穴で、彼らは若者を育て、追跡されたときにも彼らに頼ります。人が彼らに近づくと、彼らは頻繁に吠えます。彼らの音はまさに小型犬の音です。彼らはキツネと犬の中間の大きさで、非常に活発な艦隊で繊細な形をしています。耳は大きく直立し、頭を長く尖らせ、キツネのように尖らせました。話は長いです… 髪と毛皮もキツネに似ていますが、はるかに粗くて劣っています. それらは淡い赤褐色です。深海緑色の小ぶりで突き刺さる瞳。彼らのは、通常のオオカミや大西洋の州に共通するものよりもかなり長く、この地区には見られず、プラット川の上にはないと思います.
コヨーテは、1819 年 9 月、自然主義者の トーマス セイによって、スティーブン ロング少佐と政府が後援した遠征中に、プラット川の河口からミズーリ川を 24 km (15 マイル) 上流にあるルイス アンド クラークズ カウンシル ブラフスの場所で初めて科学的に記載されました。 . 彼は、1805 年 5 月 5 日付のルイスの観察のビドルの編集版を含む、ルイスとクラークのジャーナルの初版を手元に持っていました。彼の説明は 1823 年に出版されました。コヨーテ) と彼の日誌の次のページには、Canis nubilus (大平原のオオカミ) と名付けられたオオカミが掲載されています。 セイはコヨーテについて次のように説明した。
Canis latrans。灰褐色または灰色で、上部が黒色で、鈍いフルーブまたはシナモンで変化します。基部はくすんだ太い毛で、その長さの中央はくすんだシナモン色で、先端は灰色または黒色で、脊椎の方が長い。耳は直立し、先端は丸みを帯びており、後ろはシナモン色で、根元は黒く太く、内側は白髪が並んでいます。まぶたの縁は黒く、上まつげは下と上に黒い。前に黒褐色で縁取りされ、後ろに黒褐色で縁取られた補助蓋。アイリスイエロー; 瞳孔黒青; 涙嚢に黒褐色の斑点。鼻に灰色がかった色合いの演壇シナモン。唇は白く、縁取りは黒く、3 連の黒い剛毛。耳の間の頭は灰色とくすんだシナモンが混ざり合っており、毛は基部がくすんでいてふっくらしています。側面は背中よりも薄く、脚の上は時代遅れに黒く覆われています。外側の足のシナモン、後部の毛でより明確:手首の近くの前部の拡張された黒い省略線。尾はふさふさで、紡錘形で、まっすぐで、灰色とシナモンで変化し、上の基部近くの斑点があり、先端は黒です。尾の幹の先端は、足を伸ばすと、足の骨の先端に到達します。先端に向かって白く、真っ白な、テールシナモンの下に、先端は黒く。後足は 4 本指、前足は 5 本指。
命名と語源
この種に関する最初の記述は、博物学者 のフランシスコ・エルナンデスの『 Plantas y Animales de la Nueva España』(1651 年) に由来し、そこでは「スペインのキツネ」または「ジャッカル」として記述されています。「コヨーテ」という単語の最初に公開された使用法 (これは、ナワトル語の名前coyōtlのスペイン語の借用です)
![]()
発音 ( help · info ) ) は、1780 年の歴史家 FranciscoJavierClavijeroのHistoriade Méxicoします。cayjotteとcocyotieとしてさまざまに転写されます。単語のつづりは、1880 年代までに「コヨーテ」として標準化されました。コヨーテの別の英語名には、「プレーリー ウルフ」、「ブラッシュ ウルフ」、「ケースド ウルフ」、「リトル ウルフ」、「アメリカン ジャッカル」などがある。二項名のCanis latransは「吠える犬」を意味し、彼らが生み出す多くの発声に言及している。 Canis latrans の地元名と先住民名
言語グループまたは地域
土着の名前
ありから Stshirits pukatsh
カナダフランス語 コヨーテ
チヌーク
イタリパス
チペヤン Nu-ní-yĕ=̑ts!ế-lĕ
ココパ Ṭxpa Xṭpa
ノーザン クリープレインズ クリー ᒣᐢᒐᒑᑲᓂᐢ ( Mîscacâkanis ) ᒣᐢᒐᒑᑲᓂᐢ ( Mescacâkanis )
クリーク Yv•hu•ce (古風) Yv•hv•la•nu•ce (現代)
ダコタ マイカミカシカ
フラットヘッド シンクレプ
飛騨佐 モツァ
ホピ イーサウイソー
カルク ピネーフィッチ
クラマス こはあ
マンダン シェケ
マヤ語 ペキキャッシュ
ネズ・パース ʔiceyé•ye
ナワトル コヨートル
ナバホ族 マイイ
オガララ・スー Mee-yah-slay’-cha-lah
オジブウェ Mes-ta-cha’-gan-es
オマハ 三笠
オセージ ショムカシ
ポーニー チキリキ
ピウテ エジャー
スペイン語 コヨーテペロ・デ・モンテ
やかま テリパ
ティンビシャ イサ(ppü) イサパイプイサップ
ウィントゥ チャラワセデット
ヤンクトン・スー ソントケチャ
ユロク セゲプ
進化
数百万年のタイミングでのオオカミのようなイヌ科動物の系統樹
イヌ科 3.5 Ma3.0 2.5 2.0 0.96 0.6 0.38
飼い犬
![]()
灰色オオカミ
![]()
コヨーテ
![]()
アフリカオオカミ
![]()
ゴールデンジャッカル
![]()
エチオピアのオオカミ
![]()
ドール
![]()
アフリカの野生の犬
![]()
2.6
シマジャッカル
![]()
セグロジャッカル
![]()
化石記録
参照:オオカミの進化 § 化石記録

更新世のコヨーテの骨格( C. l. orcutti )
Xiaomiing WangとRichard H. Tedfordは、肉食動物の進化に関する第一人者の 1 人であり 、 Canis属はコヨーテに似たEucyon davisiの子孫であり、その化石は600万年前の中新世に最初に出現したと提案した (Mya) 。アメリカ南西部とメキシコで。鮮新世(5 Mya) までに、より大きなCanis lepophagus が同じ地域に出現し、更新世初期(1 Mya)までにC. latrans (コヨーテ) が存在していた。彼らは、Eucyon davisiからC. lepophagus、そしてコヨーテへの進化は線形進化であると提案しました。さらに、C. latransとC. aureusはC. edwardiiと密接に関連しており、ブランカン中期(鮮新世後期) からアービントン紀の終わり (更新世後期)にかけて最も早く出現した種であり、コヨーテはコヨーテと見分けがつかないままです。 C. latransは、北米でC. edwardiiと同時代でした。 Johnston は、C. lepophagusが現代のコヨーテよりも細い頭骨と骨格を持っていると説明している。 Ronald Nowak は、初期の個体群が小さなコヨーテに似ており、C. latrans .
C. lepophagusの体重は現生のコヨーテに似ていましたが、手足の骨が短く、移動の少ない生活様式を示していました。コヨーテは、オオカミが専門とする大きな獲物を保持するために必要な把持力を欠いている、その比較的小さなサイズと比較的狭い頭蓋骨と顎によって示されるように、灰色のオオカミよりも原始的なカニスの形を表しています. これは、コヨーテの矢状稜によってさらに裏付けられます。矢状稜は低いか、完全に平らであり、オオカミよりも噛みつきが弱いことを示しています。コヨーテはオオカミのように特殊な肉食動物ではなく、大臼歯の咀嚼面が大きいことからもわかるように、この種が植物に相対的に依存していることを反映しています。これらの点で、コヨーテはオオカミよりも属のキツネのような祖先に似ています。
現代のコヨーテの範囲内にある最も古い化石は、ウェスト バージニア州ハミルトン洞窟で 0.74 ~ 0.85 Ma (100 万年) のものです。0.73 Ma、カリフォルニア州アービントン。コロラド州ポーキュパイン洞窟とペンシルバニア州カンバーランド洞窟で 0.35–0.48 Ma。現代のコヨーテは、第四紀の絶滅イベントの 1,000 年後に発生しました。現代の完新世のコヨーテと比較して、更新世のコヨーテ( C. l. orcutti ) はより大きく、より丈夫であり、より大きな競争相手や獲物に反応した可能性が高い。更新世のコヨーテは、その子孫よりも特殊な肉食動物であった可能性が高く、その歯は肉の剪断により適応しており、植物を処理するのに適したすりつぶす面が少ないことが示された. 彼らのサイズの縮小は、第四紀の絶滅イベントから 1,000 年以内に発生し、大型の獲物が死にました。さらに、更新世のコヨーテはダイアオオカミ( Aenocyon dirus )の絶滅後に空いていた大物狩猟ニッチを利用することができなかった.現代の優雅なモーフを支持する自然淘汰。
DNA証拠
1993年、北米のオオカミはユーラシアのオオカミよりもコヨーテに似た頭蓋骨の特徴を示すという研究が提案されました. 2010年の研究では、コヨーテは中国チベットオオカミ、飼い犬、モンゴルオオカミ、ユーラシアオオカミを含むクレードの基本的なメンバーであり、中国チベットオオカミはオオカミや飼い犬から早期に分岐した.
2016 年に全ゲノムDNA 研究が行われた仮説に基づいて、北アメリカのオオカミとコヨーテのすべてが約 51,000 年前に共通の祖先から分岐したことが提案されました。 しかし、提案されたオオカミとコヨーテの分岐時期は、100万年前の地層でコヨーテに似た標本が発見されたことと矛盾する。この研究はまた、すべての北アメリカのオオカミにはかなりの量のコヨーテの祖先があり、すべてのコヨーテにはある程度のオオカミの祖先があり、アカオオカミと東部のオオカミはさまざまな割合の灰色のオオカミとコヨーテの祖先と高度に混合されている.
オオカミやイヌに関する遺伝子研究では、利用可能な唯一の参照ゲノムであるボクサー犬のゲノムに基づいて系統関係が推測されています。2017 年には、オオカミCanis lupus lupusの最初の参照ゲノムがマッピングされ、将来の研究に役立てられました。 2018 年に、オオカミの参照ゲノムに対する核ゲノム配列の最大のデータセットをマッピングした全範囲の標本を使用して、北アメリカのオオカミ、オオカミのようなイヌ科動物、およびコヨーテのゲノム構造と混合物を調べた研究が行われました。
この研究は、北米のハイイロオオカミとオオカミに似たイヌ科動物が複雑なハイイロオオカミとコヨーテの混合の結果であるという以前の研究結果を支持しています。グリーンランドのホッキョクオオカミとメキシコのコヨーテは、最も純粋な標本を表しています。アラスカ、カリフォルニア、アラバマ、ケベックのコヨーテには、オオカミの祖先はほとんど見られません。ミズーリ州、イリノイ州、フロリダ州のコヨーテは、オオカミの祖先の 5 ~ 10% を示します。アカオオカミでは 40% のオオカミから 60% のコヨーテの祖先があり、イースタン ティンバー オオカミでは 60% のオオカミから 40% のコヨーテ、五大湖のオオカミでは 75% のオオカミから 25% のコヨーテがありました。メキシコのオオカミと大西洋岸のオオカミではコヨーテの祖先が 10%、太平洋岸とイエローストーンのオオカミでは 5%、カナダ列島のオオカミでは 3% 未満でした。3 番目のイヌ科動物が北米のオオカミに似たイヌ科動物の混合に関与していた場合、その遺伝的特徴はコヨーテとオオカミで発見されたはずですが、発見され
2018 年には、イヌ属のメンバーを比較するために全ゲノム シーケンスが使用されました。この研究は、コヨーテとハイイロオオカミの共通の祖先が、絶滅した正体不明のイヌ科のゴースト個体群と遺伝的に混ざり合ったことを示しています。「ゴースト」イヌ科動物は遺伝的にドールに近く、アフリカの野生の犬が他のイヌ科動物種から分岐した後に進化した. オオカミと比較したコヨーテの基底位置は、コヨーテが未知の絶滅したイヌ科動物からのミトコンドリアゲノムをより多く保持しているためであると提案されています。
亜種
2005年現在、19の亜種が認められています。 コヨーテの地理的変異は大きくはないが、全体として見ると、東部の亜種 ( C. l. thamnosとC. l. frustor ) は大きく、色が濃く、徐々に色が薄くなる動物である.西と北に向かってサイズが縮小し ( C. l. texensis、C. l. latrans、C. l. lestes、およびC. l. incolatus )、太平洋に向かって「黄土色」のトーン (濃いオレンジ色または茶色) が明るくなります。海岸 ( C. l. ochropus、C. l. umpquensis )、 Aridoamerica ( C. l. microdon、C. l. mearnsi )のサイズの縮小、およびメキシコと中央アメリカの暗い赤みがかった色と短い銃口への一般的な傾向人口。
亜種
三項権威 説明と画像
範囲
プレーンズ コヨーテC. l. latransは亜種を指定します

セイ、1823 [syn: C. l. nebracensis (Merriam, 1898) C. l. 淡蒼球(メリアム、1897)]

最大の亜種。それはかなり薄い毛皮を持ち、大きな大臼歯と肉食動物を持っています。
アルバータ州、マニトバ州、サスカチュワン州南部からニューメキシコ州、テキサス州パンハンドルに至る大平原
メキシカン コヨーテC. l. カゴッティ

CEH スミス、1839 年
C.l.に似ています。半島ですが、より大きく、色が赤くなっています。耳が短く、歯が大きく、銃口が広い。
メキシコのオアハカ州、サン ルイス ポトシ州、プエブラ州、ベラクルス州
サン ペドロ マルティール コヨーテC. l. クレプティカス

エリオット、1903
小型の亜種で、夏の赤みを帯びた毛皮と、短く幅の広い頭骨を持っています。
バハ・カリフォルニア北部とカリフォルニア南西部
エルサルバドル コヨーテC. l. ディケイ

ネルソン、1932
大きな亜種で、C. l. に匹敵します。サイズは小さいですが、歯が小さく、毛皮が暗くなっています。
元々は、エルサルバドルのラ ウニオンにあるゴアスコラン川の西 3.2 km (2 マイル) にあるセロ モゴテからのみ知られていた。 2013 年 1 月、生息域を南のパナマ南部まで拡大した。
サウスイースタン コヨーテC. l. 欲求不満

ウッドハウス、1851
この亜種はC. l.に似ています。半島、しかし大きくて薄く、耳が短く、銃口が長い.
カンザス州南東部および最東端、オクラホマ州、テキサス州、ミズーリ州、アーカンソー州
ベリーズ コヨーテC. l. ゴールドマニ

メリアム、1904
メキシコのコヨーテの中で最大のもので、C. l. に近づきます。ラトランスのサイズですが、銃口が短くなっています。
グアテマラ西部のコヨーテである可能性がありますが、グアテマラ国境近くのメキシコ、チアパス州サン ビセンテでのみ知られています。
ホンジュラス コヨーテC. l. ホンジュレンシス

ゴールドマン、1936
小さくて荒っぽい色の亜種で、粗くて薄い毛皮と広い頭骨を持っています。
テグシガルパの北、アルカガの北東の開けた国でのみ知られている
デュランゴ コヨーテC. l. インパビダス

アレン、1903
このイヌ科はC. l.に似ています。カゴッティの色ですが、はるかに大きいです。
ソノラ南部、チワワ最南西部、ドゥランゴ西部、サカテカス西部、シナロア
ノーザンコヨーテC.l. インコラトゥス

ホール、1934
中型の亜種で、シナモン色の毛皮とC. l. よりも凹んだ頭骨を持っています。ラトランス。
アラスカ、ユーコン、ノースウェスト準州、ブリティッシュコロンビア州北部、アルバータ州北部の北方林
ティブロン島のコヨーテC. l. ジャメシ

タウンゼント、1912
C. l.よりもはるかに薄い。mearnsi、それはより重い歯、大きな頭蓋骨、そして長い耳を持っています.
ティブロン島
マウンテン コヨーテC. l. レステス

メリアム、1897
サイズと色がC. l. に似ています。latrans、この亜種には大きな尾と耳が
ブリティッシュ コロンビア州南部とアルバータ州南東部、ワシントン州カスケード山脈の東、オレゴン州、カリフォルニア州北部、モンタナ州西部、ワイオミング州、コロラド州(南東隅を除く)、ネバダ州北部、ユタ州北部
Mearns のコヨーテC. l. メーンシ

メリアム、1897 [syn: C. l. エスター(メリアム、1897)]

中型の耳、小さな頭蓋骨、小さな歯を持つ小さな亜種。その毛皮は豊かで明るい色です。豊かな色合いは非常に明るく、後足と前足を覆っています。
コロラド州南西部、ユタ州とネバダ州の最南端、カリフォルニア州南東部、アリゾナ州バハカリフォルニア州北東部、ニューメキシコ州のリオグランデ川の西、ソノラ州とチワワ州北部
ローワー リオ グランデ コヨーテC. l. マイクロドン

メリアム、1897
小型の亜種で、歯が小さく、毛皮はやや暗い。後足の上面は白っぽく、腹には黒い先端の毛が散りばめられています。
テキサス南部とタマウリパス北部
カリフォルニア バレー コヨーテC. l. オクロプス

フォン・エシュショルツ、1829年
C.l.に似ています。latransとC.l. lestes、しかし、より小さく、より暗く、より明るい色。耳が大きく、頭蓋骨と歯が小さい。
シエラネバダ山脈の西にあるカリフォルニア
ペニンシュラ コヨーテC. l. 半島

メリアム、1897
C.l.に似ています。サイズと機能はオクロプスですが、毛皮はより暗く、より赤くなっています。尾の下側はC. l.よりも黒くなっています。オクロプス、腹には黒い先端の毛が
バハ・カリフォルニア
イースタン コヨーテC. l. 変数。
ローレンスとボサート、1969 [syn: C. l. オリエンス、C.l.バージンシス]

これはC. lupusとC. latransのハイブリッドです。東部のオオカミよりも小さく、より小さな領土を保持しますが、典型的な西部のコヨーテよりも大きく、より広範な行動圏を保持します.
ニューイングランド、ニューヨーク、ニュージャージー、ペンシルバニア、オハイオ、ウェストバージニア、メリーランド、デラウェア、バージニア、カナダ東部のオンタリオ、ケベック、ニューブランズウィック、ノバスコシア、プリンスエドワード島、ニューファンドランドとラブラドール
テキサス プレーンズ コヨーテC. l. テキセンシス

ベイリー、1905
C.lよりも小さい。latrans、それはC. l . ocropusですが、C. l. texensisにはその亜種の大きな耳がありません。
テキサス州のほとんど、ニューメキシコ州東部、メキシコ北東部
北東コヨーテC. l. タムノス

ジャクソン、1949年
C.l.とほぼ同じサイズ。latrans、またはそれよりも大きく、色が濃く、より広い頭蓋骨を持っています。
マニトバ州サスカチュワン州中北部(南西端を除く)、ケベック州東部から南部、ノースダコタ州南部から東部、ミネソタ州、アイオワ州、ミズーリ州(ミズーリ川の北)、ミシガン州、ウィスコンシン州、イリノイ州(最南部を除く) 、およびインディアナ州北部
ノースウェスト コースト コヨーテC. l. umpquensis

ジャクソン、1949年
小型の亜種で、黒っぽい赤毛色の毛皮、比較的小さな頭蓋骨、弱い歯列を持っています。
ブリティッシュ コロンビア州、ワシントン州、オレゴン州の海岸
コリマ コヨーテC. l. 警戒

メリアム、1897
C.l.に似ています。半島ですが、より暗く、より広範囲に着色されています。前腕に黒が多く、尾の下側には黒がありません (先端を除く)。
ハリスコ南からゲレロまでのメキシコの太平洋岸
ハイブリダイゼーション

黒色のコヨーテは、飼い犬で最初に生じた突然変異に起因する色をしています。
コヨーテは飼い犬と時折交配し、口語的に「コイドッグ」として知られる交配を行うことも犬とコヨーテの交配周期は一致せず、コヨーテは通常イヌに対して敵対的であるため、このような交配は野生ではまれである. ハイブリダイゼーションは通常、コヨーテが同種がほとんどいない地域に拡大しているときにのみ発生し、犬が唯一の選択肢です. それでも、犬はコヨーテとつがいを形成しないため、子犬の生存率は通常よりも低く、子犬の飼育がより困難になります。飼育下では、F 1雑種(第 1 世代) は犬よりもいたずら好きで、子犬として扱いにくい傾向があり、オオカミと犬の雑種よりも成熟すると信頼性が低くなります . 雑種は外見が異なるが、一般的にコヨーテの通常の特徴を保持している. F 1雑種は、イヌとコヨーテの中間の形態になる傾向がありますが、F 2雑種 (第 2 世代) はより多様です。F 1ハイブリッドと F 2ハイブリッドの両方は、恥ずかしがり屋で性的攻撃性という点で、コヨーテの親に似ています。 雑種は繁殖力があり、4 世代にわたって繁殖に成功する。 黒色のコヨーテの黒い毛皮は、飼い犬で最初に生じた突然変異によるものである。ニューファンドランドの非アルビノ ホワイト コヨーテの個体群は、ゴールデン レトリバーから受け継がれたメラノコルチン 1 受容体の変異による色をしている。

オスの灰色オオカミとメスのコヨーテの間の飼育下で
生まれたコイウルフハイブリッド
コヨーテは、特に北米東部で、さまざまな程度でオオカミと交配しています。北アメリカ北東部のいわゆる「イースタン コヨーテ」は、おそらく北東部で灰色オオカミとイースタン オオカミが絶滅した後、コヨーテが以前のオオカミの生息地に定着し、生き残ったオオカミの個体群と混ざり合ったことが原因であると考えられています。この交配種は、ハイイロオオカミやイースタン オオカミよりも小さく、より小さな縄張りを保持していますが、典型的な西部のコヨーテよりも大きく、より広範な行動範囲を保持しています。2010年現在、東部のコヨーテの遺伝子構成はかなり均一で、東部のオオカミや西部のコヨーテからの影響は最小限です。大人の東部コヨーテは西部コヨーテよりも大きく、雌の東部コヨーテは雄の西部コヨーテより 21% 体重が多い。 体格の違いは生後 35 日までに明らかになり、東のコヨーテの子は西のコヨーテよりも足が長くなります。歯の発育にも違いがあり、東部コヨーテでは歯の萌出が遅く、順序が異なります。そのサイズは別として、東のコヨーテは物理的に西のコヨーテに似ています。4 つの色の段階は、暗褐色から金髪または赤みがかった金髪までの範囲ですが、最も一般的な段階は灰褐色で、脚、耳、および側面が赤みを帯びています。東部のコヨーテと西部のコヨーテの間には、攻撃性と戦闘に大きな違いはありませんが、東部のコヨーテはあまり戦う傾向がなく、より遊び心が喧嘩が遊びの行動に先行する西部のコヨーテの子犬とは異なり、東部のコヨーテの子犬の間での喧嘩は、遊びが始まった後に起こります。東部のコヨーテは、西部のコヨーテよりもはるかに遅く、2 歳で性的成熟に達する傾向が
東部オオカミとアカオオカミも、さまざまな程度のオオカミとコヨーテの交配の産物です。東部のオオカミはおそらく、オオカミとコヨーテの混血と、親のハイイロオオカミの個体群との広範な戻し交配の結果でした。アカオオカミは、南東部のウッドランズでオオカミの個体数が減少していた時期に発生した可能性があり、オオカミとコヨーテの交配が強制され、現在のアカオオカミのゲノムの約 75 ~ 80% が地域の親コヨーテの個体群と戻し交配された可能性がコヨーテ由来。
行動
社会的および生殖行動

Mearns’ coyote ( C. l. mearnsi ) の子犬が遊んでい
ます
イエローストーン国立公園のコヨーテの群れ
ユーラシアのイヌジャッカルのように、コヨーテは群生しますが、オオカミのようなより社会的なイヌ科の種ほど同種に依存しこれは、コヨーテが後者の種のように大きな獲物を狩る専門のハンターではないためと考えられます。コヨーテの群れの基本的な社会的単位は、繁殖するメスを含む家族である。ただし、無関係のコヨーテは、交際のために力を合わせたり、単独で攻撃するには大きすぎる獲物を倒したりすることがこのような「家族以外」の群れは一時的なものにすぎず、独身のオス、生殖能力のないメス、成体以下の若者で構成されている可能性がメスが発情期に入ると、真冬に家族が形成されます。つがいは、実際の交尾が行われる 2 ~ 3 か月前に起こります。交尾の結びつきは 5 ~ 45 分続くことが発情期に入ったメスは、香りのマーキングと遠吠えによってオスを惹きつける。 1 頭の雌が発情すると、最大 7 頭の生殖可能な雄を引き付けることができ、雌は 1 か月も雌を追いかけることができます。オス同士で喧嘩が起こることもありますが、メスが交尾相手を選んで交尾すると、拒絶されたオスは介入せず、他の発情したメスを見つけると動き出します。オオカミは一夫一婦制と重婚の両方の交配を行うことが知られている とは異なり、コヨーテの密集度が高く、食物が豊富な地域であっても、コヨーテは厳密に一夫一婦制です。交尾に失敗した雌は、姉妹や母親が子犬を育てるのを手伝ったり、次に交尾できるようになるまで兄弟に加わったりすることがあります. 次に、新しく交配したつがいは領土を確立し、独自の巣穴を構築するか、放棄されたアナグマ、マーモット、またはスカンクの地球を一掃します. 妊娠中、オスは頻繁に単独で狩りをし、メスのために食べ物を持ち帰ります。女性は、乾いた草または腹から引き抜かれた毛皮で巣穴を裏打ちする場合が妊娠期間は 63 日で、平均の産卵数は 6 であるが、その数はコヨーテの個体数密度と食物の豊富さによって変動する.
コヨーテの子犬は、巣穴、中空の木、または棚の下で生まれ、出生時の体重は 200 ~ 500 g (0.44 ~ 1.10 ポンド) です。彼らは晩成性であり、最初の 10 日間は牛乳に完全に依存しています。切歯は生後約 12 日、犬歯は生後16 日、第 2小臼歯は生後 21 日で萌出します。10 日後に目が開き、その時点までに子犬はますます動きやすくなり、20 日歩けるようになり、6 週齢で走るようになります。 . 両親は 12 ~ 15 日後に逆流した固形食で子犬の食事を補い始めます。生後 4 ~ 6 週間で、乳歯が完全に機能するようになると、マウス、ウサギ、または有蹄動物の死骸などの小さな餌が与えられ、 2 か月後には授乳が徐々に減少します。オオカミの子犬とは異なり、コヨーテの子犬は、遊びの行動に従事する前に、(ごっこ遊びとは対照的に) 真剣に戦い始めます。一般的な遊びの行動には、コヨーテの「ヒップスラム」が含まれます。生後 3 週間までに、コヨーテの子はオオカミの子よりも抑制が少なく互いに噛み合います。生後 4 ~ 5 週間で、子犬は優勢な階層を確立し、それまでに戦うよりも遊ぶ可能性が高くなります。オスは子犬に餌をやったり、グルーミングしたり、守ったりするのに積極的な役割を果たしますが、子犬が完全に離乳する前にメスが行方不明になると、それらを捨てます. 巣穴は6月から7月まで放棄され、子犬は親に続いて縄張りを巡回し、狩りをします。子犬は 8 月に家族のもとを離れることもありますが、ずっと長く留まることができます。子犬は生後 8 か月で成体の大きさになり、1 か月後には成体の体重が増えます。
縄張り意識と保護行動
個々の摂食領域は、0.4 から 62 km2 (0.15 から 24 平方マイル) の範囲でサイズが異なります。特定の地域におけるコヨーテの一般的な濃度は、食物の豊富さ、適切な巣穴の場所、および同種や他の捕食者との競争によって異なります。コヨーテは一般に、巣を作る季節以外は自分の縄張りを守らず、オオカミよりも侵入者に対して攻撃的ではなく、通常、侵入者を追いかけたりスパーリングしたりしますが、殺すことはめったにありません。コヨーテ間の対立は、食糧不足の時期に発生する可能性がコヨーテは足を上げて排尿し、地面を引っ掻いて自分の縄張りをマークします。
オオカミのように、コヨーテは巣穴 (通常は他の種のさびれた穴) を使用して妊娠し、若い子を育てますが、屋外のヤマヨモギの下で出産することもコヨーテの巣窟は、峡谷、ウォッシュアウト、クーリー、バンク、岩の絶壁、または平地に位置する可能性が放棄されたホームステッド小屋、穀物貯蔵庫、排水管、線路、中空の丸太、茂み、およびアザミの下にいくつかの巣穴が見つかりました。巣穴は、子犬が生まれるまで、メスによって継続的に掘られ、掃除されます。巣穴が乱されたり、ノミがはびこったりした場合、子犬は別の巣穴に移されます。コヨーテの巣穴には、メインチャンバーから分岐するいくつかの入り口と通路が 1 つの巣穴は毎年使用できます。
狩猟と摂食行動
嗅覚は狩猟にとって非常に重要であるというのが一般的なコンセンサスであるが、嗅覚、聴覚、および視覚的な合図の役割を実験的に調査した 2 つの研究では、アカギツネとコヨーテの狩猟には視覚的な合図が最も重要であることがわかった。 .

獲物に飛びかかるコヨーテ。
大きな獲物を狩るとき、コヨーテはペアまたは小さなグループで行動することがよく大型の有蹄動物を殺す成功は、雪の深さや地殻の密度などの要因に左右されます。若い動物は通常、そのような狩猟に参加することを避け、通常、繁殖ペアがほとんどの作業を行います。コヨーテは大きな獲物を追いかけ、典型的にはハムストリングを行い、獲物が倒れるまで嫌がらせをする. 他のイヌ科動物と同様に、コヨーテは余分な食物を蓄えます。コヨーテはネズミほどの大きさのげっ歯類を飛び跳ねて捕まえるが、ジリスは追いかける。コヨーテは大きな群れで生活することができますが、小さな獲物は通常単独で捕獲されます。コヨーテはペアでヤマアラシを殺し、足を使ってげっ歯類を背中にひっくり返し、柔らかい下腹部を攻撃することが観察されています。年配の経験豊富なコヨーテだけがヤマアラシを首尾よく捕食できます。コヨーテは、おそらく食べ物の所有権を主張するために、食べ物に排尿することが 最近の証拠によると、少なくとも一部のコヨーテは、おそらく人間を避けるために、狩猟においてより夜行性になっている. コヨーテは時折、アメリカアナグマと相利共生の狩猟関係を形成し、げっ歯類の獲物を掘り起こす際に互いに助け合うことがある. 2 種の関係は時折、明らかに「友情」に近いものになる場合があり、一部のコヨーテはアナグマの仲間の上に頭を横たえたり、抗議せずに顔をなめたりしているのが観察されています。コヨーテとアナグマの間の友好的な相互作用は、コロンブス以前の文明に知られており、メキシコで見つかった 1250 年から 1300 年のCEの瓶に示されているように、両者の関係が描かれています。
生ゴミ、ペットフード、動物の糞は、コヨーテをゴミ箱に引き寄せる可能性が
コミュニケーション

コヨーテの遠吠え
夜に吠えるコヨーテの群れ
ボディランゲージ
群生と孤独の両方の動物であるため、コヨーテの視覚と発声のレパートリーの多様性は、孤独なキツネと非常に社交的なオオカミの中間です。コヨーテの攻撃的な行動は、オオカミやイヌよりもキツネに似ている. 攻撃的なコヨーテは背中を丸め、尻尾を下げます。「プレイボウ」に続いて「プレイリープ」を実行することで遊び心のある行動を求める犬とは異なり、コヨーテの遊びは、弓とそれに続く左右の頭の屈曲と一連の「スピン」で構成されます。 」と「ダイブ」。コヨーテは犬のように遊び仲間の首筋を噛むことがありますが、通常は低く近づき、上向きに噛みつきます。子犬は性別に関係なく互いに戦いますが、大人の間では、攻撃性は通常、同性のメンバーに予約されています. 戦闘は通常、静かに行われますが、戦闘員は尻尾を振ってあごを開けて唸りながらお互いに近づきます。男性は垂直の姿勢で戦う傾向がありますが、女性は4本の足すべてで戦います。メス同士の喧嘩は、オス同士の喧嘩よりも深刻になる傾向があり、メスは相手の前足、喉、肩をつかみます。
発声
叫ぶコヨーテ
コヨーテは、「すべての 北アメリカの哺乳動物の中で最も声がよい」と言われています。 その声の大きさと発声範囲が、「吠える犬」を意味する二項名のCanis latransの原因となった。大人のコヨーテでは、少なくとも 11 の異なる発声が知られています。これらの音は、敵意と警告、挨拶、連絡の 3 つのカテゴリに分類されます。最初のカテゴリの発声には、ウーフ、うなり声、ハフ、吠え声、樹皮の遠吠え、叫び声、高頻度の泣き声が含まれます。横糸は低強度の脅威または警報として使用され、通常、巣穴の近くで聞こえ、子犬がすぐに巣穴に後退するように促します。うなり声は、近距離での脅威として使用されますが、遊んで交尾しているオスの間でも聞こえてきました。ハフは、空気の急速な呼気によって生成される高強度の脅威の発声です。鳴き声は、遠距離からの脅威の発声と警告音の両方に分類できます。樹皮の遠吠えも同様の機能を果たす可能性が叫び声は服従のしるしとして発せられますが、高頻度の鳴き声は、支配的な動物が部下の服従を認めることによって生成されます。挨拶の発声には、低周波の泣き声、「うわー、うわー」、およびグループのイップハウリングが含まれます。低周波の鳴き声は従順な動物が発するもので、通常は尻尾を振ったり銃口をかじったりします。「うわーうわー」として知られる音は、「挨拶の歌」と表現されています。グループ イップの遠吠えは、2 人以上のパック メンバーが再会したときに発せられ、複雑な挨拶式の最後の行為である可能性が連絡先の通話には、単独の遠吠えとグループの遠吠え、および前述のグループのイップの遠吠えが含まれます。孤独な遠吠えはコヨーテの最も象徴的な音であり、群れから離れた孤独な個体の存在を知らせる目的に役立つ可能性がグループの遠吠えは、代わりのグループのイップの遠吠えとして、および単独の遠吠え、グループの遠吠え、またはグループのイップの遠吠えへの応答として使用されます。
エコロジー編集
生息地

サンフランシスコのバーナルハイツにある都会のコヨーテ
オオカミとクーガーがほぼ絶滅する前に、コヨーテはバイソン、プロングホーン、ヘラジカ、その他のシカが生息する草原に最も多く生息し、プレーリードッグのいる短い草地で特によく暮らしていました。よもぎやジャックラビットが生息する半乾燥地帯、またはサボテン、カンガルーネズミ、ガラガラヘビが生息する砂漠。オオカミと直接競合しない限り、コヨーテはソノラ砂漠から隣接する山々の高山地域、またはアルバータ州の平野と山岳地帯にまで及びました。オオカミの絶滅により、コヨーテの範囲はグアテマラの熱帯地方とアラスカの北斜面からの壊れた森林を含むように拡大しました.
コヨーテは 1 日に約 5 ~ 16 キロメートル (3 ~ 10 マイル) 歩きます。彼らは冬の間、氷に覆われた川を移動ルートとして使用することが彼らはしばしば薄明薄暮性であり、日中よりも夕方と夜の始まりに活動的です. 多くのイヌ科動物と同様に、コヨーテは有能な水泳選手であり、少なくとも 0.8 キロメートル (0.5 マイル) 水面を移動できると報告されています。
ダイエット

ワイオミング州シーズカディー国立野生生物保護区で、道路で殺されたプロングホーンの切れ端を持つコヨーテ
コヨーテは、生態学的にはユーラシアのイヌジャッカルに相当する北米の動物です。同様に、コヨーテは食物の選択において非常に用途が広いが、主に肉食であり、その食事の 90% は肉から成っている. 獲物の種には、バイソン(主に腐肉)、オジロジカ、ミュールジカ、ヘラジカ、ヘラジカ、オオツノヒツジ、プロングホーン、ウサギ、ノウサギ、げっ歯類、鳥(特にガリギズ、ロードランナー、若い水鳥、ハト、ハト)、両生類が含まれます(ヒキガエルを除く)、トカゲ、ヘビ、カメ、カメ、魚、甲殻類、昆虫。トガリネズミ、モグラ、ドブネズミなどの動物は、その数に比例して食事に含まれないため、コヨーテはターゲットとする獲物にうるさい場合がしかし、ジリスや関連種 (マーモット、プレーリードッグ、シマリス)などの陸生および/または穴を掘る小型哺乳類、ハタネズミ、ポケットホリネズミ、カンガルーラット、およびその他の地面を好むげっ歯類は、特に一般的な食物である可能性が孤独なコヨーテのために。 より珍しい獲物には、漁師、若いツキノワグマの子、ハープ アザラシ、ガラガラヘビなどがコヨーテは、ガラガラヘビが伸びるまでヘビをからかってから、頭を噛んだり、ヘビをパチンと振ったりすることで、主に食べ物のために(しかし、巣穴で子犬を保護するために)ガラガラヘビを殺します。コヨーテに捕らえられる鳥のサイズは、スラッシャー、ヒバリ、スズメから大人の野生の七面鳥、まれに成体の白鳥やペリカンまでさまざまです。 群れやつがいで行動する場合、コヨーテは、体重が 10 kg (22 ポンド) を超えるさまざまな獲物など、単独の個体が通常取るよりも大きな獲物にアクセスできる可能性が 場合によっては、コヨーテの群れがオドコイレウスの成獣、ウシのヘラジカ、プロングホーン、野生のヒツジなどのはるかに大きな獲物を送り出しているが、これらの動物の若い子鹿、子牛、子羊は、かなり頻繁に捕食されている。パック、国産羊、国産牛。場合によっては、コヨーテは体重が 100 ~ 200 kg (220 ~ 440 ポンド) 以上の獲物を倒すことができます。野生のシカなどの成体の有蹄動物に関して言えば、弱っていたり、雪や氷に閉じ込められていたり、冬に衰弱していたり、重度の妊娠をしていたりするなど、脆弱なときに搾取されることがよくありますが、用心深い家畜の有蹄動物はより簡単に搾取される可能性があります.
コヨーテは新鮮な肉を好みますが、機会があればあさります。食べる昆虫、果物、草を除いて、コヨーテは毎日推定600g(1.3ポンド)、または年間250kg(550ポンド)の食物を必要とします. コヨーテは同種の死骸を容易に共食いし、コヨーテ ハンターはコヨーテの脂肪をルアーや毒餌として首尾よく使用した 。コヨーテの冬の食事は、主に大きな有蹄動物の死体で構成され、植物はほとんど含まれげっ歯類の獲物は、春、夏、秋に重要性が増します。
コヨーテは、ブラックベリー、ブルーベリー、桃、ナシ、リンゴ、ウチワサボテン、シャポテ、柿、ピーナッツ、スイカ、マスクメロン、ニンジンなど、さまざまな農産物を食べます。冬から早春にかけて、コヨーテは緑の小麦の葉などの草を大量に食べます。綿菓子、大豆粕、家畜の糞、豆、トウモロコシ、小麦、モロコシなどの栽培穀物などの珍しいものを食べることも
カリフォルニアの沿岸では、コヨーテは祖先よりも海洋性食品を多く消費しています。これは、この地域からハイイログマが絶滅したためと考えられています。デスバレーでは、コヨーテは春の開花期にスズメガの 幼虫やカブトムシを大量に消費することがある.
敵と競争相手

コヨーテとオオカミの比較イラスト

若いクーガーを追い詰めるマウンテン コヨーテ ( C. l. lestes )
コヨーテとハイイロオオカミの生息域が重なっている地域では、干渉競争とオオカミによる捕食が、地域のコヨーテ密度を制限していると仮定されています。オオカミが絶滅した後、19 世紀から 20 世紀にかけてコヨーテの生息域が拡大しましたが、1940 年代にオオカミが島に定着した後、アイル ロイヤルではコヨーテが絶滅に追い込まれました。両方の種が共存するイエローストーン国立公園で実施されたある研究では、1990 年代にオオカミが再導入された後、ラマー リバー バレーのコヨーテの個体数は39 %減少したと結論付けられました。ない地域よりも33%低くなります。 オオカミは近くのコヨーテを容認しないことが観察されているが、コヨーテはオオカミを追跡して獲物を食べることが知られている.
コヨーテは、一部の地域でクーガーと競合する場合がシエラネバダ東部では、コヨーテがミュールジカをめぐってクーガーと競い合います。クーガーは通常、コヨーテを打ち負かして支配し、時折コヨーテを殺すことが殺されたコヨーテは食べられないこともあり、これはおそらくこれらが競争的な種間相互作用を構成していることを示しているが、クーガーがコヨーテも食べていることが確認された事例が複数ある. メキシコ北東部では、コヨーテに対するクーガーの捕食が急速に続いているが、明らかに生息地の用途が異なるため、コヨーテは同所ジャガーの獲物スペクトルには含まれていなかった.
オオカミやクーガー以外では、大人のコヨーテを捕食することは比較的まれですが、他の複数の捕食者が時折脅威になる可能性があります. 場合によっては、成体のコヨーテがアメリカクロクマとグリズリーベア 、アメリカアリゲーター、カナダオオヤマネコ、イヌワシ の両方に捕食されている. 殺害現場や腐肉では、コヨーテは、特に単独で行動する場合、オオカミ、クーガー、クマ、クズリ、そして常にではないが通常はワシ(すなわち、禿げたものや金色のもの)に支配される傾向がある。これらのようなより大きく、より強力で、より攻撃的な捕食者が共有の餌場に来ると、コヨーテは戦おうとするか、他の捕食者が終わるまで待つか、時折殺しを共有するかもしれませんが、オオカミや大人のクーガーがいると、コヨーテは逃げる傾向が
コヨーテは、健康なアカギツネの成体を殺すことはめったになく、わなに引っかかったキツネを殺すことがよくありますが、コヨーテと一緒に餌を食べたり巣穴を掘ったりすることが観察されています。コヨーテはキツネのキットを殺すかもしれませんが、これは主要な死因ではありません. 南カリフォルニアでは、コヨーテは頻繁にハイイロギツネを殺し、これらの小さなイヌ科動物は、コヨーテの密度が高い地域を避ける傾向が
一部の地域では、コヨーテはボブキャットと範囲を共有しています。ボブキャットの個体数は、コヨーテの密度が高い地域では減少する傾向がありますが、これら 2 つの同じようなサイズの種が物理的に衝突することはめったにありません。しかし、いくつかの研究は、コヨーテとボブキャットの間の干渉競合を示しており、すべてのケースでコヨーテが相互作用を支配していました。 複数の研究者 は、コヨーテがボブキャットを殺した例を報告したが、ボブキャットがコヨーテを殺した例はよりまれである。コヨーテは、中型の獲物に使用されるものと同様に、噛んで振る方法を使用してボブキャットを攻撃します。コヨーテ (単独の個人とグループの両方) は、時折ボブキャットを殺すことが知られています。ほとんどの場合、ボブキャットは成体の雌や幼体などの比較的小さな個体でした。しかし、大人のオスのボブキャットに対するコヨーテの攻撃 (不明な数のコヨーテによる) が発生しています。カリフォルニアでは、コヨーテとボブキャットの個体数は異なる生息地タイプ間で負の相関はありませんが、コヨーテによる捕食はボブキャットの死亡率の重要な原因です. 生物学者のスタンリー・ポール・ヤングは、捕獲したボブキャットがコヨーテに殺されるのを防いだことは一度もないと指摘し、コヨーテがボブキャットを追いかけて木の上に登った 2 つの事件について書いている。コヨーテは時々カナダ オオヤマネコを直接殺し 、獲物、特にカンジキウサギをめぐって彼らと競争することが記録されている。アルバータ州中部を含むいくつかの地域では、オオヤマネコはコヨーテが少ない場所でより豊富に生息しているため、コヨーテとの相互作用は、カンジキウサギの入手可能性よりもオオヤマネコの個体数に影響を与えるように思われる.
範囲
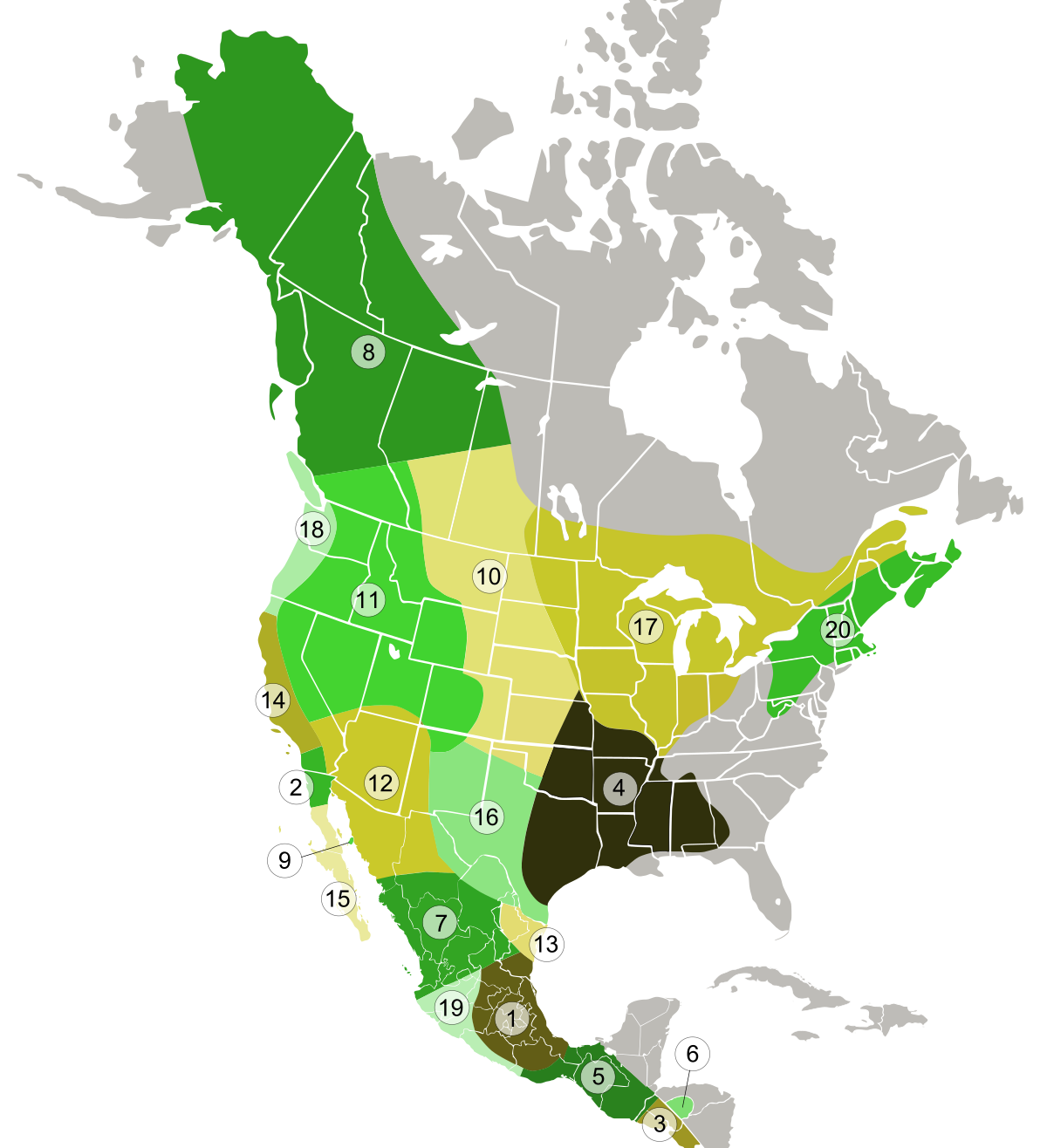
1978年現在のコヨーテ亜種の範囲: (1) メキシココヨーテ、(2) サンペドロ・マルティールコヨーテ、(3) エルサルバドルコヨーテ、(4) 南東部コヨーテ、(5) ベリーズコヨーテ、(6) ホンジュラスコヨーテ、(7)デュランゴ コヨーテ、(8) ノーザン コヨーテ、(9)ティブロン島コヨーテ、(10)平野コヨーテ、(11) マウンテン コヨーテ、(12)ミーンズ コヨーテ、(13) ローワー リオ グランデ コヨーテ、(14) カリフォルニア バレー コヨーテ、 (15) ペニンシュラ コヨーテ、(16) テキサス プレーンズ コヨーテ、(17) 北東コヨーテ、(18) 北西海岸コヨーテ、(19) コリマ コヨーテ、(20)イースタン コヨーテ

過去1万年間のコヨーテの拡大

1900年以来の数十年にわたるコヨーテの拡大
コヨーテは北米全体に広く生息しているため、国際自然保護連合(IUCN)によって軽度懸念事項に指定されています。コロンブス以前のコヨーテの生息域は、北米の南西部と平原、およびメキシコ北部と中央部に限定されていた。19 世紀までにこの種は北と東に拡大し、1900 年以降はさらに拡大し、土地の転換とオオカミの絶滅と一致しました。この時までに、その範囲は、隣接するすべての米国とメキシコ、南は中央アメリカ、北はカナダとアラスカの大部分を含む北米大陸全体を網羅していました。この拡大は進行中であり、この種は現在、北緯 8 度 (パナマ) から北緯 70 度 (アラスカ北部) の間の地域の大部分を占めています。
コヨーテはメキシコ南部と中央アメリカへの最近の移民であると広く信じられていましたが、更新世と完新世初期の記録、およびコロンブス以前と初期のヨーロッパの植民地時代の記録は、この動物が森林伐採によって拡大したことを示しています近世よりずっと前からこの地域に存在する。それにもかかわらず、1970 年代後半にはコスタリカ南部で、1980 年代前半にはパナマ北部で生息域の拡大が起こりました。コヨーテは近い将来ベリーズ北部に出現すると予測されている。ダリエンギャップがパンアメリカンハイウェイによって閉鎖された場合、パナマ地峡を通って南アメリカに拡大する可能性があるという懸念が提起されている. この恐怖は、2013 年 1 月にパナマ運河を越えた東パナマのチェポ地区で種が記録されたときに部分的に確認された.
2017 年の遺伝子研究では、コヨーテはもともと米国東部の地域では発見されていなかったことが示唆されています。1890 年代から、密集した森林が農地に変わり、オオカミの駆除が大規模に実施され、コヨーテが分散するニッチが残されました。遺伝的に異なるコヨーテの 2 つの集団から 2 つの主要な分散がありました。北東部への最初の大規模な分散は、20 世紀初頭にグレート プレーンズ北部に生息するコヨーテからもたらされました。これらは、五大湖地域北部とカナダ南部を経由してニューイングランドに到着し、五大湖地域南部を経由してペンシルベニアに到着し、1940 年代にニューヨークとペンシルベニアで出会いました。これらのコヨーテは、コヨーテの遺伝的多様性を増し、新しいニッチへの適応を助けた可能性がある残りの灰色のオオカミとイースタンオオカミの集団と交配しました. 南東への 2 番目の主要な分散は、20 世紀半ばにテキサスから来て、1980 年代にカロライナに達しました。これらのコヨーテは、アカオオカミが野生で絶滅した 1970 年代以前に残ったアカオオカミの個体群と交配し、コヨーテの遺伝的多様性も増し、この新しいニッチへの適応にも役立った可能性がこれら 2 つの主要なコヨーテの分散は両方とも、急速な個体数の増加を経験しており、大西洋中部の海岸に沿って集まると予測されています。この研究は、コヨーテの場合、長距離分散、局所個体群からの遺伝子の流れ、および急速な個体群の増加が相互に関連している可能性があると結論付けています。
病気と寄生虫

疥癬に苦しむカリフォルニア バレー コヨーテ ( C. l. ochropus )
大型の北米の肉食動物の中で、コヨーテはおそらくその広範囲で多様な食事のために、おそらく最も多くの病気と寄生虫を運びます. コヨーテに感染することが知られているウイルス性疾患には、狂犬病、犬ジステンパー、伝染性犬肝炎、馬脳炎の 4 系統、および口腔乳頭腫症が含まれます。1970 年代後半までに、コヨーテでの深刻な狂犬病の発生は 60 年以上問題ではなくなっていましたが、1 ~ 5 年ごとに散発的な症例が発生していました。ジステンパーは、野生では多くの子犬を死に至らしめていますが、一部の個体は感染しても生き残ることができます。細菌性疾患である野兎病は、マダニに刺されたり、齧歯類やうさぎ類の獲物を介してコヨーテに感染し、子犬にとって致命的となる可能性が
コヨーテは、疥癬と疥癬の両方に感染する可能性があり、後者が最も一般的です. コヨーテではダニの侵入はまれで偶発的ですが、マダニの侵入はより一般的で、地域によって季節的なピークがあります (北西部では 5 月から 8 月、アーカンソー州では 3 月から 11月)。コヨーテにシラミが寄生することはめったにありませんが、ノミは子犬の頃からコヨーテに寄生しますが、深刻な病気よりも刺激の原因となる可能性があります. Pulex simulansはコヨーテに寄生する最も一般的な種ですが、Ctenocephalides canisは、コヨーテとイヌ (主な宿主) が同じ地域に生息する場所でのみ発生する傾向がコヨーテが吸虫の宿主になることはめったにありませんが、それでもコヨーテに深刻な影響を与える可能性があり、特にNanophyetus salmincolaは、死亡率が 90% のサケ中毒病に感染する可能性が吸虫メトルキス・コンジャンクタスもコヨーテに感染することが サナダムシは、調べたすべてのコヨーテの 60 ~ 95% に寄生することが記録されています。コヨーテに寄生する最も一般的な種は、ワタオウサギを中間宿主として使用するTaenia pisiformisとTaenia crassicepsです。コヨーテで知られている最大の種はT. hydatigenaで、感染した有蹄動物からコヨーテに入り、80 から 400 cm (31 から 157 インチ) の長さに成長することができます。かつては主にオオカミに限定されていましたが、コヨーテが以前のオオカミの生息地に定着し始めて以来、エキノコックス グラニューロサスはコヨーテにまで拡大しました。コヨーテで最も頻繁に発生する回虫はToxascaris leoninaで、これはコヨーテの小腸に生息し、宿主がより頻繁に食事をすることを除いて、悪影響はありません。アンシロストマ属の鉤虫は、その範囲全体でコヨーテに寄生し、特に湿気の多い地域で流行しています。テキサス沿岸などの湿気の多い地域では、コヨーテはそれぞれ最大 250 匹の鉤虫を運ぶことができます。血を飲むA. caninumは、失血と肺のうっ血によってコヨーテにダメージを与えるため、特に危険です。10 日齢の子犬は、わずか 25匹のA. caninumワームの宿主であるために死亡する可能性が
人間関係
詳細情報:アーバン コヨーテ
民話や神話では
コヨーテ (神話)

エドワード・S・カーティスの昔のインディアン時代にカヌーで漕ぐコヨーテ
コヨーテは、一部のネイティブ アメリカン、特に南西部および平原地域のいくつかの国で、実際のコヨーテまたは人間の形を交互にとる民話のトリックスターフィギュアおよびスキン ウォーカーとして登場します。他のトリックスターのフィギュアと同様に、コヨーテは欺瞞とユーモアを通じて社会的慣習に反抗するピカレスクなヒーローとして行動します。ハリスなどの民俗学者は、動物の知性と適応性のために、コヨーテがトリックスターと見なされるようになったと信じています。ヨーロッパによるアメリカ大陸の植民地化の後、英米のコヨーテの描写は、臆病で信用できない動物である. コヨーテに対する英米の文化的態度は、大衆的なイメージが根本的に改善されたハイイロオオカミとは異なり、大部分が否定的なままである.
マイドゥの創造の物語で、コヨーテは世界に仕事、苦しみ、そして死をもたらします。ズニ族の伝承では、コヨーテがカチナから光を盗むことで世界に冬をもたらしています。チヌーク、マイドゥ、ポーニー、トホノ オッダム、ウテは、コヨーテを創造主の仲間として描いています。Tohono O’odham の洪水の物語では、人類を滅ぼす世界的な大洪水からモンテスマが生き残るのをコヨーテが助けています。クリエイターが人類を創造した後、コヨーテとモンテスマは人々に生き方を教えます。クロウの創造物語では、コヨーテ老人が創造主として描かれています。The Dineh Creation Storyでは、コヨーテは最初の男性と最初の女性と共に第一世界に存在していましたが、別のバージョンでは第四世界で作成されています. ナバホ コヨーテは世界に死をもたらし、死がなければあまりにも多くの人々が存在し、トウモロコシを植える余地がなくなると説明しています。

コヨーテの戦士を描いた
テオティワカンのアテテルコの壁画
スペインがアステカ帝国を征服する前、コヨーテはメソアメリカの宇宙論において重要な役割を果たしていました。コヨーテは古典時代の テオティワカンの軍事力を象徴し、戦士はコヨーテの衣装を着てその略奪力を呼び起こしました. この種は、ポスト古典アステカの支配に至るまでの何世紀にもわたって、中央メキシコの戦士のカルトに結び付けられ続けました. アステカ神話では、ダンス、音楽、肉欲の神であるフエウエコトル(「年老いたコヨーテ」を意味する) が、コヨーテの頭を持つ男としていくつかの写本に描かれている。彼は時々女たらしとして描かれ、愛の女神であるソチケツァルを誘惑することで世界に戦争をもたらした. エピグラファーの デビッド・H・ケリーは、ケツァルコアトル神は、人類の「兄」、創造者、誘惑者、詐欺師、文化の英雄として描かれているコヨーテのアステカ以前のユート・アステカ神話の描写にその起源を負っていると主張した。明けの明星へ。
人間への攻撃
詳細は「コヨーテの人間への攻撃」を
参照

人々がコヨーテに餌を与えることを思いとどまらせる兆候。これにより、コヨーテは人間の存在に慣れ、攻撃の可能性が高まります。
コヨーテの人間への攻撃はまれであり、コヨーテのサイズが比較的小さいため、重傷を負うことはめったにありませんが、特にカリフォルニアではますます頻繁になっています. 致命的な攻撃が確認されたのは 2 つだけです。1 つはカリフォルニア州グレンデールの3 歳のケリー キーンで、もう 1 つはカナダのノバスコシア州のテイラー ミッチェルという 19 歳の少年です。 2006 年 3 月までの 30 年間で、米国では少なくとも 160 件の攻撃が発生し、主にロサンゼルス郡地域で発生しました。米国農務省 (USDA) 野生生物局、カリフォルニア州魚類狩猟局、およびその他の情報源からのデータは、1988 年から 1997 年の間に 41 件の攻撃が発生した一方で、1998 年から 2003 年にかけて 48 件の攻撃が検証されたことを示しています。これらの事件の大部分は、郊外と野生地の境界に近い南カリフォルニアで発生しました。
農村部の人々によるコヨーテへの嫌がらせがなければ、都会のコヨーテは人間への恐怖を失いつつあり、意図的または無意識にコヨーテに餌を与える人々によってさらに悪化しています. このような状況では、一部のコヨーテは人間に対して攻撃的に行動し始め、ジョギングをしている人や自転車を追いかけたり、犬の散歩をしている人々に立ち向かったり、小さな子供をストーカーしたりしています. これらの地域の非狂犬病のコヨーテは、ほとんどが 10 歳未満の小さな子供を標的にすることがあるが、噛まれた大人もいる.
そのような攻撃のメディア報道は、一般的に問題の動物を単に「コヨーテ」と特定していますが、コヨーテ東部の遺伝学に関する研究は、ペンシルバニア、ニューヨーク、ニューイングランド、およびカナダ東部を含む北アメリカ北東部での攻撃に関与している可能性があることを示しています。実際にはコイウルフ、Canis latransとC. lupusのハイブリッドであり、完全なコヨーテではありません。
家畜とペットの捕食

犬と対峙するコヨーテ
2007年現在、コヨーテは北アメリカ西部で最も豊富な家畜の捕食者であり、羊、山羊、牛の大部分が失われました. たとえば、国家農業統計局によると、 2004年に捕食が原因で死亡した 224,000 頭の羊の 60.5% がコヨーテによるものでした。米国の羊と子羊の総個体数の 2.22% であり、全米農業統計局 USDA の報告によると、2005 年 7 月 1 日時点でそれぞれ合計 466 万頭と 780 万頭であるコヨーテの個体数は通常、コヨーテはオオカミよりも何倍も大きく、より広く分布していますが、コヨーテはより多くの全体的な捕食損失を引き起こします. 米国政府機関は、家畜を保護するために、毎年約 90,000 匹のコヨーテを日常的に撃ち、毒を与え、罠にかけ、殺しています。 2005 年に行われたアイダホ州の国勢調査では、個々のコヨーテが個々のオオカミよりも家畜を攻撃する可能性が 5% 高いことが示されました。ユタ州では、2017 年 6 月 30 日に終了した会計年度に合計 50 万ドル以上の懸賞金のために 11,000 匹以上のコヨーテが殺されました。
家畜の保護犬は、捕食者を積極的に撃退するために一般的に使用されており、フェンスで囲まれた牧草地と放牧地の両方でうまく機能しています。 1986 年の米国の羊生産者の調査では、82% が犬の使用が経済的資産であると報告していることがわかりました。
コロラド州立大学のテンプル・グランディンが論じたコヨーテを制御する方法として、牛の自然な保護傾向を高めることを含む牛の再野生化がこの方法は、牧草地で牛の分娩を許可し、牛が年間を通して放牧地を放牧する生産者の間で人気を集めています。

飼いならされた羊に典型的な喉のホールドを持つコヨーテ
コヨーテは成体のヒツジやヤギを攻撃する際に、通常、顎のすぐ後ろと耳の下の喉を噛み、窒息死することが多い. 失血は通常、二次的な死因です。子牛や毛の多い羊は、側面や後肢を攻撃して殺され、ショックや失血を引き起こします。若い子羊などの小さな獲物を攻撃する場合、頭蓋骨と脊椎の領域を噛むことで殺し、組織と骨に大きな損傷を与えます. 小さな獲物や若い獲物は完全に運び去られ、殺害の証拠として血だけが残ります。コヨーテは通常、大型動物の皮と骨格の大部分を比較的無傷のまま残しますが、餌が不足している場合は、最大の骨のみを残すことがコヨーテがより大きな死体を広範囲に食べている場合、羊毛、皮、およびその他の部分が散らばっているのが特徴です。
足跡は、コヨーテと犬の捕食を区別する重要な要素です。コヨーテの足跡は、飼い犬よりも楕円形でコンパクトな傾向があり、犬の足跡よりも爪跡が目立たず、直線に近い傾向がサイトハウンドを除いて、コヨーテと同程度の体重のほとんどの犬は歩幅がわずかに短くなっています。 コヨーテの殺害は、オオカミの殺害とは、下にある組織への損傷が少ないことで区別できます。また、コヨーテの糞はオオカミの糞よりも小さい傾向が
コヨーテは、獲物として現れるほど小さいドッグフードや動物に惹かれることがよくゴミ、ペットフード、時には鳥やリスの餌場などのアイテムは、コヨーテを裏庭に引き寄せます. コヨーテに攻撃された約 3 ~ 5 匹のペットが毎週、南オレンジ郡(カリフォルニア州)のアニマル アージェント ケア病院に運ばれています。猫は通常攻撃に耐えられないため、その大部分は犬です。カリフォルニア州クレアモントの近くで収集されたスカトロ分析により、コヨーテは冬と春の食料源としてペットに大きく依存していることが明らかになりました。南カリフォルニアのある場所で、コヨーテは食料源として野良猫のコロニーに依存し始めた. 時間が経つにつれて、コヨーテはほとんどの猫を殺し、猫のコロニーを維持していた人々がコロニーのサイトに毎日配置したキャットフードを食べ続けました. コヨーテは通常、小型の犬を攻撃しますが、例外的なケースでは、ロットワイラーなどの大型で強力な犬種でさえも攻撃することが知られています. グレイハウンドなどのコヨーテよりも大きな犬は、一般的にそれらを追い払うことができ、コヨーテを殺すことが知られています. 小型の品種は、怪我や死亡に苦しむ可能性が高くなります。
狩猟
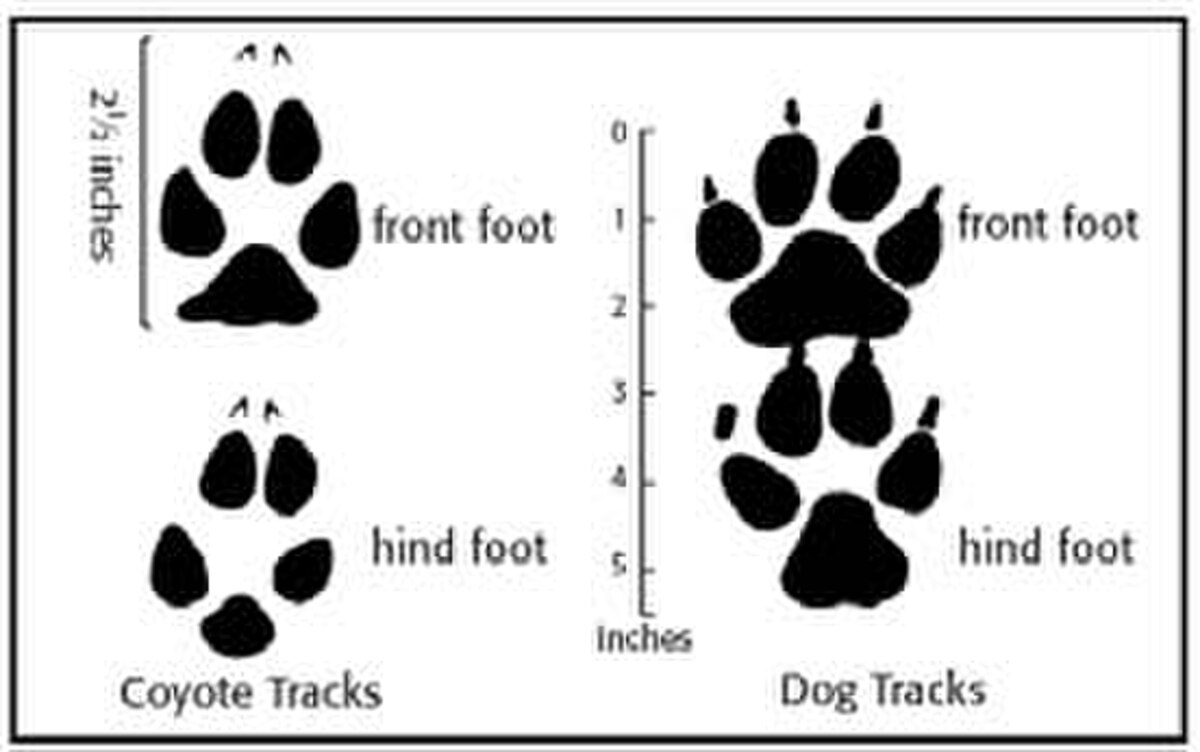
飼い犬の足跡と比較したコヨーテの足跡
コヨーテの狩猟は、人間が参加する捕食者の狩猟の最も一般的な形態の 1 つです。コヨーテの捕獲に関して多くの規制はありません。つまり、動物を狩るために使用できるさまざまな方法が最も一般的な形式は、トラッピング、コーリング、ハウンドハンティングです。コヨーテは色覚異常であるため、灰色と微妙な青の色合いでしか見ることができず、迷彩や無地のパターンを使用することができます. 平均的なオスのコヨーテの体重は 8 ~ 20 kg (18 ~ 44 ポンド) で、平均的なメスのコヨーテの体重は 7 ~ 18 kg (15 ~ 40 ポンド) であるため、これらの重量の間で実行できる汎用発射体は.223 レミントンであり、発射体はエントリー後、エグジット前にターゲット内で膨張するため、最も多くのエネルギーが供給されます。コヨーテは軽くて機敏な動物であるため、地形に非常に軽い印象を与えることがよくコヨーテの足跡は長方形で、長さ約 6.35 cm (2.5 インチ)、幅 5.08 cm (2 インチ) です。前足と後足の両方に4本の爪がコヨーテのセンター パッドは、比較的丸みを帯びた三角形のような形をしています。飼い犬と同様に、コヨーテの前足は後足よりわずかに大きいです。コヨーテの足は飼い犬の足に最も似ています。
毛皮の用途

カナダのコヨーテの毛皮
19 世紀半ばまで、コヨーテの毛皮は価値がないと考えられていました。これはビーバーの減少とともに変化し、1860 年までに毛皮を目的としたコヨーテの狩猟は、大平原のオオカミにとって大きな収入源 (1 皮あたり 75セントから1.50ドル) になりました。コヨーテの毛皮は、1950 年代初頭に経済的に重要な存在であり、価格は産地に応じて 1 枚あたり 5 ドルから 25 ドルでした。コヨーテの毛皮はラグを作るには十分な耐久性がありませんが、コートやジャケット、スカーフ、またはマフに使用できます. 毛皮の大部分は、婦人服のコートの襟や袖などのトリミングに使用されます。コヨーテ の 毛皮 は 、シルバー フォックスの 模造 品 と し て 黒く 染め られる こと も あ る。
西部開拓時代、コヨーテは時々わな猟師や山岳民族に食べられました。コヨーテは平原インディアンの饗宴に登場することがあり、カリフォルニア州サンガブリエルの先住民はコヨーテの子犬を食べていました。コヨーテの肉の味はオオカミの味に例えられ、茹でると豚肉よりも柔らかくなります。コヨーテの脂肪は、秋に摂取すると、革にグリースを塗ったり、スプレッドとして食べたりすることがあります.
飼いならしやすさ
コヨーテは、コロンブス以前のさまざまな文化によって半家畜化された可能性が19 世紀の作家の中には、コヨーテがグレート プレーンズの原住民の村で飼われていると書いたものがコヨーテは子犬の頃は簡単に飼いならすことができますが、大人になると破壊的になることが純血コヨーテと雑種コヨーテはどちらも、遊び心があり、所有者に打ち明けることができますが、懐疑的で見知らぬ人には恥ずかしがり屋ですが 、コヨーテは回収や指差しなどの実用的な目的に使用するのに十分扱いやすいですが、記録されます。 1945 年の夏に捕獲された「ブッチ」という名前の飼いならされたコヨーテは、映画でのキャリアは短命で、鶏小屋を襲撃しているときに撃たれる前に、スモーキー(1946) とラムロッド(1947) に出演しました。
有名メディアで
ワイリー E. コヨーテは、短編アニメ映画のルーニー テューンズとメリー メロディーズシリーズで際立って登場します。
アリゾナ州のNHLチームは、この地域にコヨーテが多数生息していることに敬意を表し、アリゾナ・コヨーテと名付けられました。
The Good, The Bad and The Ugly (1966) で有名なoo-wee-oo-wee-oo wah-wah-wah の叫び声は、コヨーテの遠吠えに触発されました。
ノート
^ 「ケース入りオオカミ」という名前は、歴史的にコヨーテの皮膚がマスクラットのようにケースに入れられていたのに対し、オオカミはビーバーのように平らに広げられていたことに由来する. ^ 参考文献の完全なセットについては、オオカミの進化#オオカミのようなイヌ科の系統樹の注記 (a) を参照して
引用
^ Tedford, Wang & Taylor 2009 , p. 131。
^ Kays, R. (2020) 。「カニス・ラトランス」。絶滅危惧種の IUCN レッド リスト。2018 : e.T3745A163508579. doi : 10.2305/IUCN.UK.2018-2.RLTS.T3745A163508579.en . 2021年11 月 11 日閲覧。
^
ジェームズ、エドウィン。ロング、スティーブン・H。ねえ、トーマス。アダムス、ジョン(1823)。1819 年と 20 年に行われた、ピッツバーグからロッキー山脈への遠征の記録。巻。1. ロンドン:ロングマン、ハースト、ピーズ、オーレ、ブラウン。pp.168–174。
^ 「カニス・ラトランス」。Fossilworks.org . 2021年12 月 17 日閲覧。
^
ベコフ M. (1977)。「カニス・ラトランス」。哺乳類の種(79): 1–9. ドイ: 10.2307/3503817 . ISSN 1545-1410。JSTOR 3503817 . OCLC 46381503。
^ ヤング & ジャクソン 1978 年、p. 48 ^ Young & Jackson 1978 , pp. 63–4
^ ヤング & ジャクソン 1978 , pp. 50–53 ^ ヤング & ジャクソン 1978 年、p. 247
^ フォックス 1978 年、 p. 105
^ 「オオカミと土地を共有する」 (PDF) . ウィスコンシン州天然資源局。2015 . 2016年6 月 29 日閲覧。
^ Cartaino 2011、p。16 ^ ヤング & ジャクソン 1978 年、p. 59
^ ヴァンタッセル、スティーブン(2012)。「コヨーテ」。鳥獣被害調査ハンドブック(第3版)。ネブラスカ州リンカーン: 野生生物管理コンサルタント。p。112.ISBN _ 978-0-9668582-5-9. OCLC 794471798。
^ Nowak 1979、p。14.
^ ホフマイスター、ドナルド F. (2002)。イリノイ州の哺乳類。イリノイ大学プレス。pp.33–34。ISBN 978-0-252-07083-9. OCLC 50649299。
^ Mussulman、Joseph 。「コヨーテ」。ルイス&クラークを発見。2013年1 月 15 日閲覧。
^ Mussulman、Joseph 。「トーマス・セイ:Canis latrans」 . ルイス&クラークを発見。2013 年 7 月 21 日にオリジナルからアーカイブされました。2013年1 月 15 日閲覧。
^ クラビジェロ、フランシスコ・ハビエル。カレン、チャールズ(1817)。メキシコの歴史: スペイン人とメキシコ人の歴史家から、インディアンの写本と古代の絵画から集められたもの: スペイン人によるメキシコの征服とともに: メキシコの土地、動物、住民に関する重要な論文を含む彫刻によって説明されています. 巻。1.フィラデルフィア:トーマス・ドブソン。p。57. OCLC 13601464 .
^ Bullock、W.(1824)。メキシコでの 6 か月間の滞在と旅行: ヌエバ スペインの現在の状態、その自然生産物、社会の状態、製造業、貿易、農業、古代などについての発言が含まれています。: プレートと地図付き。ロンドン: ジョン・マレー、アルベマール通り。119ページ、261ページ。
^ Seton 1909、p。789
^ k l
Gier、HT (1974)。「コヨーテ ( Canis latrans )の生態と行動」. In Fox, MW (ed.)。野生のイヌ科:その系統、行動生態学、および進化。ニューヨーク:ヴァン・ノストランド・ラインホールド。pp.247–262。ISBN 978-0-442-22430-1. OCLC 1093505 . ^ g
ベコフ、マーク。Gese、Eric M.(2003)。「コヨーテ」。Feldhamer、George A.; トンプソン、ブルースC。チャップマン、ジョセフ A. (eds.)。北米の野生哺乳類: 生物学、管理、および保全(第 2 版)。メリーランド州ボルチモア: ジョンズ・ホプキンス大学出版局。pp.467–470。ISBN 978-0-8018-7416-1. OCLC 51969059。
^ Lehner、Philip N. (1978)。「コヨーテ通信」。In Bekoff, M. (ed.)。コヨーテ: 生物学、行動、および管理。ニューヨーク:アカデミックプレス。pp.127–162。ISBN 978-1-930665-42-2. OCLC 52626838。
^ k Young & Jackson 1978 , pp. 6–7
^ カーティス、ES(1928)。チペヤン。西部の森のクリー。サルシ。北米インディアン。巻。18.クラシックブックスカンパニー。p。201.ISBN _ 978-0-7426-9818-5.
^ クロフォード、JM(1989)。ココパ辞典。カリフォルニア大学出版局。p。445.ISBN _ 978-0-520-09749-0. OCLC 20012309 .
^ ルクレア、N.Cardinal, G. (1998)。アルバータ長老のクリー辞書。アルバータ大学。p。279.ISBN _ 978-0-88864-284-4. OCLC 659111819。
^ マーティン、JP; モールディン、MM (2004)。クリーク/マスコギーの辞書。ネブラスカ大学出版局。p。153.ISBN _ 978-0-8032-8302-2. OCLC 43561668。
^ アルバート、R.; ショール、DL (1985)。簡潔なホピと英語の辞書。ジョン・ベンジャミンズ・パブリッシング. p。26.ISBN _ 978-90-272-2015-8. OCLC 777549431。
^ ブライト、ウィリアム。ゲール、スーザン。「コヨーテ」。Karuk辞書とテキスト。Karuk Tribe & UC Berkeley . 2015年5 月 22 日閲覧。
^
リード、FA(2009)。中央アメリカとメキシコ南東部の哺乳類のフィールド ガイド。オックスフォード大学出版局。p。259.ISBN _ 978-0-19-534322-9. OCLC 237402526。
^ 青木治夫(1994).ネズ・ペルセ辞書。カリフォルニア大学出版局。p。491.ISBN _ 978-0-520-09763-6. OCLC 463788185。
^ Neundorf、A.(1983)。ナバホ族/英語のバイリンガル辞書: Áłchíní Bi Naaltsoostsoh . ニューメキシコ大学出版局。p。512.ISBN _ 978-0-8263-3825-9. OCLC 57357517。
^ Quintero、C.(2004)。オーセージ文法。ネブラスカ大学出版局。p。83.ISBN _ 978-0-8032-3803-9. OCLC 57614396。
^ パークス、RP。プラット、LN (2008)。Skiri Pawnee の辞書。ネブラスカ大学出版局。p。119.ISBN _ 978-0-8032-1926-7. OCLC 940905155。
^
Dayley、JP(1989)。Tümpisa (Panamint) ショショーニ語辞典。カリフォルニア大学出版局。p。436.ISBN _ 978-0-520-09754-4. OCLC 489876664。
^ Pitkin, H. (1985)。ウィントゥ辞書。カリフォルニア大学出版局。pp. 65, 573.ISBN 978-0-520-09613-4. OCLC 12313411。
^ 「コヨーテ」。ユロク辞書:セゲプ。ユロク言語プロジェクト。UC バークレー校。2015年5 月 22 日閲覧。
^ 「自然史:家族の絆」 . アメリカ自然史博物館の雑誌。巻。117、いいえ。6. ニューヨーク:アメリカ自然史博物館。2008年。22.
^ 「Canis lepophagus」。化石工場。2021年12 月 17 日閲覧。
^ Wang & Tedford 2008、p. 58. ^ Tedford, Wang & Taylor 2009 , pp. 175, 180. ^ ジョンストン 1938 年、p. 385。
^ Nowak 2003、p。241。
^ Nowak、RM(1978)。「コヨーテと関連するCanisの進化と分類」. In Bekoff, M. (ed.)。コヨーテ: 生物学、行動、および管理。ニューヨーク:アカデミックプレス。pp. 3–16。ISBN 978-1-930665-42-2. OCLC 52626838。
^ Tedford, Wang & Taylor 2009 , p. 136。
^
ジャワ州ミーチェン; サミュエルズ、JX (2012)。「メガファウナルの絶滅に対応したコヨーテ(Canis latrans )の進化」 . 米国科学アカデミーの議事録。109 (11): 4191–6. ビブコード: 2012PNAS..109.4191M . ドイ: 10.1073/pnas.1113788109 . ISSN 1091-6490。OCLC 475396714。PMC 3306717 . PMID 22371581。
^ ジャワ州ミーチェン; ジャノウィッツ、AC; エイブリー、JE。サドレア、RW (2014)。「氷河期のメガファウナル絶滅に対応したコヨーテ(Canis latrans )の生態学的変化」 . プロスワン。9 (12): e116041. ビブコード: 2014PLoSO…9k6041M . ドイ: 10.1371/journal.pone.0116041 . PMC 4281224 . PMID 25551387。
^ Goulet、GD (1993)。近極期、現代、完新世、更新世後期のハイイロオオカミ (Canis lupus) と選択された Canis (修士論文) の間の頭蓋骨の時間的および地理的変異の比較。ウィニペグにあるマニトバ大学。pp. 1–116。
^ チャン、ホンハイ。チェン、レイ (2010)。「ドール Cuon alpinus の完全なミトコンドリアゲノム: イヌ科内の系統発生分析と年代測定の進化的分岐」. 分子生物学レポート。38 (3): 1651–60. ドイ: 10.1007/s11033-010-0276-y . PMID 20859694。S2CID 7476436 . ^
ボンホールド、BM。ケイヒル、ジャワ; ファン、Z。グロナウ、I.; ロビンソン、J。ポーリンジャー、JP。シャピロ、B。ウォール、J。ウェイン、RK(2016)。「全ゲノム配列解析により、北米オオカミの 2 つの固有種がコヨーテとハイイロオオカミの混合物であることが示されました」 . 科学の進歩。2 (7): e1501714。Bibcode : 2016SciA….2E1714V . ドイ: 10.1126/sciadv.1501714 . PMC 5919777 . PMID 29713682。
^ バージニア州モレル (2016)。「オオカミじゃないオオカミをどうやって救うの?」科学。353 (6300)。ドイ: 10.1126/science.aag0699 . ^ Wang & Tedford 2008、p.
^ ゴパラクリシュナン、シャム。サマニエゴ カストルイータ、ホセ A.; Singing、Mikkel-Holger S.; クデルナ、ルーカス FK。ライコネン、ヤニッケ; ピーターセン、ベント。Sicheritz-Ponten、トーマス。ラーソン、グレガー。オーランド、ルドビッチ。マルケス・ボネ、トーマス。ハンセン、アンダース J.; ダレン、愛。ギルバート、M. トーマス P. (2017)。「オオカミの参照ゲノム配列 (Canis lupus lupus) とその Canis spp. 集団ゲノミクスへの影響」 . BMCゲノミクス。18 (1): 495. doi : 10.1186/s12864-017-3883-3 . PMC 5492679 . PMID 28662691。
^ Singing、Mikkel-Holger S.; ゴパラクリシャン、シャム。Vieira、Filipe G.; サマニエゴ カストルイータ、ホセ A.; ラウンドラップ、カトリーヌ。ハイデ・ヨルゲンセン、マッツ・ピーター。Meldgaard、Morten。ピーターセン、ベント。Sicheritz-Ponten、トーマス。ミケルセン、ヨハン・ブルース。マーカルド・ピーターセン、ウルフ。ディーツ、ルーン; ゾンネ、クリスチャン。ダレン、愛。バッハマン、ルッツ。ウィグ、オイシュタイン; ハンセン、アンダース J.; ギルバート、M. トーマス P. (2018)。「北米における灰色オオカミとオオカミのようなイヌ科動物の集団ゲノミクス」 . PLOS遺伝学。14 (11)。e1007745。ドイ: 10.1371/journal.pgen.1007745 . PMC 6231604 . PMID 30419012。
^ ゴパラクリシュナン、シャム。Singing、Mikkel-Holger S.; ラモス・マドリガル、ジャズミン。ニーマン、ジョナス。サマニエゴ カストルイータ、ホセ A.; Vieira、Filipe G.; カロエ、クリスチャン。モンテロ、マルク・デ・マヌエル。クデルナ、ルーカス。セレス、アイトール。ゴンザレス・バサロテ、ビクトル・マヌエル。劉燕胡。王、国東; マルケス・ボネ、トーマス。Mirarab、Siavash。フェルナンデス、カルロス。ゴーベール、フィリップ。Koepfli、Klaus-Peter。バッド、ジェーン。Rueness、Eli Knispel。ハイデ・ヨルゲンセン、マッツ・ピーター。ピーターセン、ベント。Sicheritz-Ponten、トーマス。バッハマン、ルッツ。ウィグ、オイシュタイン; ハンセン、アンダース J.; ギルバート、M. トーマス P. (2018)。「イヌ属の進化を形作った種間遺伝子流動」 . 現在の生物学。28 (21): 3441–3449.e5. ドイ: 10.1016/j.cub.2018.08.041 . PMC 6224481 . PMID 30344120。
^ Wozencraft、WC(2005)。「食肉目」 . デラウェア州ウィルソン。Reeder, DM (eds.)。世界の哺乳類種: 分類学的および地理的参照(第 3 版)。ジョンズ・ホプキンス大学出版局。pp.532–628。ISBN 978-0-8018-8221-0. OCLC 62265494。
^ ヤング & ジャクソン 1978 年、p. 249 ^ g h i
メリアム、CH (1897)。「コヨーテまたはプレーリーオオカミの改訂、新しい形態の説明」 . ワシントン生物学会の議事録。11 :19~33。
^ r s Nowak
1979 , pp. 9–10
^ エリオット、DG(1904)。中米と西インド諸島の陸と海の哺乳類。巻。Ⅱ.イリノイ州シカゴ。pp.467–468。
^ ネルソン、EW(1932)。「サルバドールからの新しい亜種の説明を含む、コヨーテに関するコメント」 . ワシントン生物学会の議事録。45 : 223–225. 2014 年 9 月 7 日にオリジナルからアーカイブされました。2014年9 月 7 日閲覧。
^ Méndez-Carvajal、P. (2014)。「哺乳類、肉食動物、イヌ科、イヌラトランス(Say, 1823): パナマでの実際の分布」 . チェックリスト。10 (2): 376–379. ドイ: 10.15560/10.2.376 . ISSN 1809-127X . OCLC 828104819 。2015年1 月 26 日閲覧。
^ メリアム、CH (1904)。「メキシコ南部からの新しいコヨーテ」 . ワシントン生物学会の議事録。17 : 157. 2014 年 9 月 7 日に元の場所からアーカイブされました。
^ ゴールドマン、EA (1936)。「ホンジュラスから来た新しいコヨーテ」 . J.Wash.Acad.科学。26:32~34。
^ ヤング & ジャクソン 1978 年、p. 263
^ タウンゼント、CH (1912)。「1911 年にローワー カリフォルニアで「アルバトロス」探検隊によって収集された哺乳動物と、新種の説明」 . アメリカ自然史博物館紀要。31 : 117–130.
^ ベイリー、V. (1905)。「テキサスの生物調査」 . 北米の動物相。25 : 1 ~ 222。ドイ: 10.3996/nafa.25.0001 . hdl : 2027/mdp.39015006867405 . ISSN 1944-4575 . OCLC 273060038。
^ ジャクソン、HHT (1949)。「アメリカから来た2匹の新しいコヨーテ」 . ワシントン生物学会の議事録。62 : 31–32.
^ アンダーソン、TM。ボンホールド、BM。Candille、SI; Musiani、M。グレコ、C.スターラー、DR; スミス、DW。パドゥカサハスラム、B.; ランディ、E。レナード、JA; ブスタマンテ、CD; Ostrander、EA; タン、H.; ウェイン、RK; Barsh、GS(2009)。「北米ハイイロオオカミにおけるメラニズムの分子および進化史」 . 科学。323 (5919): 1339–1343. Bibcode : 2009Sci…323.1339A . ドイ: 10.1126/science.1165448 . ISSN 1095-9203。OCLC 34298537。PMC 2903542 . PMID 19197024。
^ Young & Jackson 1978 , pp. 121–24 ^ Cartaino 2011 , pp. 61–3
^ フォックス 1978 年、 p. 136
^ ジマー、カール (2013 年 1 月 21 日)。「スノーコヨーテとスピリットベア」 . ナショナル ジオグラフィック マガジン。2016年7 月 1 日閲覧。
^
ウェイ、JG。ラトリッジ、L.; ウィールドン、T.; BN ホワイト (2010)。「マサチューセッツ州東部の東部「コヨーテ」の遺伝的特性」 (PDF) . 東北ナチュラリスト。17 (2): 189–204. ドイ: 10.1656/045.017.0202 . ISSN 1938-5307 . JSTOR 40664873 . OCLC 46381506。S2CID 135542 .
^ ウェイ、JG(2007)。「北アメリカ東部と西部のCanis latrans (コヨーテ)の体重の比較」 (PDF) . 東北ナチュラリスト。14 (1): 111–24. ドイ:10.1656/1092-6194(2007)142.0.CO;2 . ISSN 1938-5307 . JSTOR 4499900 . OCLC 46381506。S2CID 85288738 .
^ Bekoff、M.(1978)。「コヨーテとイースタンコヨーテの行動発達」. コヨーテ: 生物学、行動、および管理。ニューヨーク:アカデミックプレス。pp.97–127。ISBN 978-1-930665-42-2. OCLC 52626838。
^ ヒルトン、ヘンリー(1978)。「イースタンコヨーテの系統と生態」. In Bekoff, M. (ed.)。コヨーテ: 生物学、行動、および管理。ニューヨーク:アカデミックプレス。pp.210–28。ISBN 978-1-930665-42-2. OCLC 52626838。
^ ボンホールド、BM。ポーリンジャー、JP。アール、DA; ノウルズ、JC。アーカンソー州ボイコ; パーカー、H。ゲフェン、E。パイロット、M。Jedrzejewski、W。Jedrzejewska、B。シドロビッチ、V。グレコ、C.ランディ、E。Musiani、M。ケイズ、R。ブスタマンテ、CD; Ostrander、EA; Novembre,J.ウェイン、RK(2011)。「謎めいたオオカミのようなイヌ科動物の進化の歴史に関するゲノム全体の展望」 . ゲノム研究。21 (8): 1294–1305。ドイ: 10.1101/gr.116301.110 . ISSN 1549-5469。OCLC 37589079。PMC 3149496 . PMID 21566151。
^ Fox、MW(1974)。「イヌ科動物の社会的行動の進化」. 野生のイヌ科: その系統学、行動生態学、および進化。ニューヨーク:ヴァン・ノストランド・ラインホールド。pp.429–60。ISBN 978-0-442-22430-1. OCLC 1093505 .
^ ベコフ、マーク。ダイヤモンド、ジュディ(1976)。「コヨーテの交尾前および交尾行動」. ジャーナル オブ マンマロジー. 57 (2): 372–375. ドイ:10.2307/1379696。ISSN 0022-2372。JSTOR 1379696 . OCLC 1800234。
^ カールソン、デブラ A.; Gese、Eric M.(2008)。「コヨーテ (Canis latrans) の生殖生物学: 交尾行動、生殖ホルモン、および膣細胞学の統合」 . ジャーナル オブ マンマロジー. 89 (3): 654–664. doi : 10.1644/06-mamm-a-436r1.1 . PMC 7108653 . PMID 32287378。
^ ゲセ、エリックM。ラフ、ロバート L. (1997)。「コヨーテ、カニス・ラトランスによる香りのマーキング:社会的および生態学的要因の影響」. 動物の行動。54 (5): 1155–1166. CiteSeerX 10.1.1.540.1024 . doi : 10.1006/anbe.1997.0561 . PMID 9398369。S2CID 33603362 .
^ Mech、DL (2003)。ミネソタのオオカミ:ハートランドで遠吠え。ボイジャープレス。p。75.ISBN _ 978-0-89658-509-6. OCLC 43694482。
^ カリフォルニア州ヘネシー。Dubach、J.; Gehrt、SD(2012)。「都会のコヨーテ ( Canis latrans )における一夫一婦制の長期的なつがいと遺伝的証拠」 . ジャーナル オブ マンマロジー. 93 (3): 732–742. ドイ: 10.1644/11-MAMM-A-184.1 . ISSN 1545-1542。OCLC 39098574。
^ Fox 1978年、p. 33
^ Mlot、Chris (1998)。「ラマー バレーのコヨーテ: イエローストーンでは、マスター アダプターがオオカミの扱い方を学習します」. 科学ニュース。153 (5): 76–78. ドイ:10.2307/4010114。JSTOR 4010114 .
^ Wells、Michael C.、および Marc Bekoff. 「コヨーテ Canis latrans の匂いマーキングの観察研究」(1981)。
^ ヤング & ジャクソン 1978 , pp. 82–90
^ アサ、CS; Mech, D. (1995)。「オオカミの感覚器官とその生活史への重要性に関するレビュー」. カービン、LD。フリッツ、SH; Seip、DR(編)。変化する世界におけるオオカミの生態と保護。アルバータ州エドモントン: アルバータ大学。pp.287–291。ISBN 978-0-919058-92-7. OCLC 35162905。
^ Österholm、H.(1964)。「キツネ ( Vulpes vulpes L. )の摂食行動における距離受信の重要性」. Acta Zoologica Fennica . 106 : 1–31.
^ ウェルズ、MC (1978)。「捕食におけるコヨーテの感覚 – それらの相対的な使用に対する環境の影響」. 行動プロセス。3 (2): 149–158. ドイ:10.1016/0376-6357(78)90041-4 . PMID 24924653。S2CID 22692213 .
^ ウェルズ、MC。Lehner、PN(1978)。「コヨーテの捕食行動における距離感覚の相対的重要性」. 動物の行動。26 : 251–258. ドイ:10.1016/0003-3472(78)90025-8 . S2CID 53204333 . ^ ヤング & ジャクソン 1978 , pp. 91–92 ^ ヤング & ジャクソン 1978 年、p. 97 ^ ヤング & ジャクソン 1978 年、p. 98
^ ジェイコブス、ジュリア (2018 年 6 月 15 日)。「哺乳類は人間を避けるために夜行性になる」 . ニューヨークタイムズ。2022 年 1 月 1 日にオリジナルからアーカイブされました。2018年6 月 21 日閲覧。
^ ゲイナー (2021 年 6 月 15 日)。「野生生物の夜行性に対する人間の撹乱の影響」 . 科学。360 (6394): 1232–1235. ドイ: 10.1126/science.aar7121 . PMID 29903973 . S2CID 49212187 . 2021年11 月 14 日閲覧。
^ 米国魚類野生生物局 (2016)。「発見!一緒に狩りをするコヨーテとアナグマ」 . ^ Young & Jackson 1978 , pp. 93–96
^ 「どうしてうちの庭にコヨーテがいるの?食べ物のルアーやその他の答え」 . 米国人道協会。2020年5月7日閲覧。
^ シルバー、H。シルバー、WT(1969)。「ニューイングランド北部のコヨーテに似たイヌ科動物の成長と行動、およびイヌ科雑種の観察」. 野生生物協会、野生生物モノグラフ。17 (17): 24–25. ISSN 1938-5455 . JSTOR 3830473 . OCLC 60618095。
^ Fox 1978 , pp. 134–135
^ ビー、ジェームズ(1981)。カンザスの哺乳類。カンザス大学。p。165。
^ マイケル・A・マレス; オクラホマ自然史博物館 (オクラホマ州ノーマン) (1999). 砂漠の百科事典。オクラホマ大学出版局。pp.137–8。ISBN 978-0-8061-3146-7.
^ サンダース、DA 「アディロンダック生態センター:コヨーテ」。環境科学林業大学、SUNY。
^ ホール、ロバートL.; シャープ、ヘンリー S. (1978)。オオカミと人間:並行進化。ニューヨーク:アカデミックプレス。p。156.ISBN _ 978-0-12-319250-9. OCLC 3607816。
^ Minta, SC, Minta, KA, & Lott, DF (1992). アナグマ (Taxidea taxus) とコヨーテ (Canis latrans) の間の狩猟協会。ジャーナル オブ マンマロジー、73(4)、814–820。
^ Bartel, RA, & Knowlton, FF (2005). 1977年から1993年、ユタ州カーリューバレーでの変動する獲物量に対するコヨーテCanis latransの機能的な摂食反応。カナダ動物学ジャーナル、83(4)、569–578。
^ Best, TL, Hoditschek, B., & Thomas, HH (1981). オクラホマ州のコヨーテ (Canis latrans) の食べ物。南西部の自然主義者、26(1)、67–69。
^ ブランデージ、GC (1993)。ニューヨーク州アディロンダック中心部に生息する東部コヨーテ ( Canis latrans var.) の捕食生態(PhD)。ニューヨーク州立大学、シラキュース、環境科学および林業学部。
^ ボイヤー、RH(1949)。「セコイア国立公園でマウンテン コヨーテが 1 歳のツキノワグマを殺す」. ジャーナル オブ マンマロジー. 30 : 75. doi : 10.1093/jmammal/30.1.75 . ISSN 1545-1542。OCLC 39098574。
^ ウェイ、JG。ホートン、J. (2004)。「コヨーテがタテゴトアザラシを殺す」 (PDF) . Canidニュース。7 (1)。ISSN 1545-1542。OCLC 39098574。2006 年 5 月 13 日にオリジナル (PDF)からアーカイブされました。
^ クラウバー、ローレンス・モンロー(1997)。ガラガラヘビ:その習性、生活史、人類への影響。巻。1 (第 2 版)。カリフォルニア州バークレー: カリフォルニア大学出版局。pp.1072–1074。ISBN 978-0-520-21056-1. OCLC 39523012。
^ スペリー、CC(1939)。「ペグレッグコヨーテの食性」. ジャーナル オブ マンマロジー. 20 (2): 190–194. ドイ:10.2307/1374376。JSTOR 1374376 .
^ MacCracken、JG。Uresh、DW(1984)。「サウスダコタ州ブラックヒルズのコヨーテの食べ物」. 野生生物管理のジャーナル。48 (4): 1420–1423。ドイ:10.2307/3801809。JSTOR 3801809 . ^ スミス、JW (1988). ミズーリ州の実験的なトランペッター スワン復元プログラムの状況。手続き中。第 10 回トランペッター スワン ソサエティ会議の論文、D. コンプトン編集、100–103。ミネソタ州メープル プレイン: The Trumpeter Swan Society。
^ Bunnell, FL, Dunbar, D., Koza, L., & Ryder, G. (1981). ブリティッシュ コロンビア州のシロ ペリカンの生産性と個体数に対する撹乱の影響: 観察とモデル。植民地時代の水鳥、2–11。
^ Gese, EM, Rongstad, OJ, & Mytton, WR (1988). コロラド州南東部におけるコヨーテの群れの大きさと食事との関係。野生生物管理ジャーナル、647–653。
^ ボーエン、WD (1978). 獲物のサイズに関連するコヨーテの社会組織(博士論文、ブリティッシュ コロンビア大学)。
^ Arjo、WM; プレッチャー、DH; リーム、RR (2002)。「モンタナ州北西部におけるオオカミとコヨーテの食事の重複」 . ジャーナル オブ マンマロジー. 83 (3): 754–766. doi : 10.1644/1545-1542(2002)083<0754_dobwac>2.0.co;2 . ^ Gese, EM, & Grothe, S. (1995). ワイオミング州イエローストーン国立公園における冬のシカとヘラジカに対するコヨーテの捕食の分析。アメリカのミッドランド自然主義者、36–43。
^ Whitlaw, HA, Ballard, WB, Sabine, DL, Young, SJ, Jenkins, RA, & Forbes, GJ (1998). ニューブランズウィック州におけるオジロジカの成体の生存率と原因別死亡率。野生生物管理ジャーナル、1335-1341。
^ Bruns、EH(1970)。「プロングホーンカモシカのイヌワシとコヨーテの冬の捕食」. できる。フィールドナット。84 : 301–304. ^ ボゲス、EK、アンドリュース、RD、およびビショップ、RA (1978). アイオワ州におけるコヨーテと犬の家畜の損失。野生生物管理ジャーナル、362–372。
^ コノリー、G. (1992). 家畜やその他の資源に対するコヨーテの被害。Boer、AH、イースタン コヨーテの生態学と管理、ニュー ブランズウィック大学、ニュー ブランズウィック。
^ リード、レイチェルEB。ギフォード・ゴンザレス、ダイアン。コッホ、ポール L (2018)。「カリフォルニア沿岸におけるコヨーテ (Canis latrans) の海洋資源の利用: 彼らの最近の祖先と比較した新しい行動」 . 完新世。28 (11): 1781–1790. ビブコード: 2018Holoc..28.1781R . ドイ:10.1177/0959683618788714。
^ コーディ、ヒュー (2013)。北アメリカ: 1 つの大陸の世界。フィラデルフィア: ランニング プレス。ISBN 978-0-7624-4843-2. OCLC 808413615。
^ バーガー、KM; Gese、EM(2007)。「オオカミとの干渉競争は、コヨーテの分布と数を制限しますか?」. 動物生態学ジャーナル。76 (6): 1075–1085。ドイ: 10.1111/j.1365-2656.2007.01287.x . PMID 17922704。
^ 染色、HJ (1974)。「イヌ科の分布と分類」. In Fox, MW (ed.)。野生のイヌ科: その系統学、行動生態学、および進化。ニューヨーク:ヴァン・ノストランド・ラインホールド。pp. 3–26。ISBN 978-0-442-22430-1. OCLC 1093505 .
^ ホーノッカー、M。Negri、S.(2009)。Cougar: エコロジーと保全。シカゴ大学出版局。p。170.ISBN _ 978-0-226-35347-0. OCLC 609634655。
^ パロマレス、F。Caro, TM (1999)。「哺乳動物の肉食動物における種間殺戮」 (PDF) . アメリカのナチュラリスト。153 (5): 492–508. ドイ:10.1086/303189。hdl : 10261/51387。PMID 29578790。S2CID 4343007 . ^ Ackerman, BB, Lindzey, FG, & Hemker, TP (1984). ユタ州南部のクーガーの食習慣。野生生物管理ジャーナル、147–155。
^ エルナンデス – セント マーチン、AD。Rosas-Rosas、OC; Palacio-Núñez、J.; ルイジアナ州タランゴ アランブラ。Clemente-Sánchez、F.; Hoogesteijn、アラバマ州 (2015)。「メキシコ北東部の保護地域と隣接する断片化された景観におけるジャガーとピューマの食性」. 自然地域ジャーナル。35 (2): 308–317. ドイ:10.3375/043.035.0213。S2CID 86622145 . ^ Beer, Encyclopedia of North American Mammals: An Essential Guide to Mammals of North America . サンダーベイプレス (2004),
ISBN 978-1-59223-191-1 .
^ グギスバーグ、CAW (1972)。ワニ: その自然史、民俗学、および保全。p。195.ISBN _ 978-0-7153-5272-4. ^ O’Donoghue, M., Boutin, S., Murray, DL, Krebs, CJ, Hofer, EJ, Breitenmoser, U., Breitenmoser-Wuersten, C., Zuleta, G., Doyle, C. & Nams, VO (2001)。コヨーテとオオヤマネコ。北方林の生態系ダイナミクス: Kluane プロジェクト。オックスフォード大学出版局、ニューヨーク、ニューヨーク、米国、276–323。
^ メイソン、JR (2000)。「イヌワシがオスの成体コヨーテを攻撃して殺す」 (PDF) . 猛禽研究のジャーナル。34 (3): 244–245.
^ アレン、ML。エルブロッホ、LM; ウィルマーズ、CC; Wittmer、HU(2015)。「スカベンジャーによる腐肉の獲得に対する大型肉食動物の比較効果」. アメリカのナチュラリスト。185 (6): 822–833. ドイ:10.1086/681004。hdl : 2142/101871。PMID 25996866。S2CID 206003003 . ^ グリーン、GI、マットソン、DJ、およびピーク、JM (1997). イエローストーン国立公園のハイイログマが有蹄動物の死体を食べている春。野生生物管理ジャーナル、1040-1055。
^ ウィルマーズ、CC; スターラー、DR; クラブツリー、RL; スミス、DW。ゲッツ、WM (2003)。「資源の分散と消費者の優位性: 米国グレーター イエローストーンでオオカミとハンターに殺された死骸をあさる」 . 生態学の手紙。6 (11): 996–1003。ドイ: 10.1046/j.1461-0248.2003.00522.x . S2CID 55961335 .
^ ホーノッカー、MG。ハッシュ、HS(1981)。「モンタナ州北西部のクズリの生態」. カナダ動物学ジャーナル。59 (7): 1286–1301。ドイ:10.1139/z81-181。
^ ボーエン、WD (1980)。「有蹄動物の死体でのコヨーテとイヌワシの相互作用」. J.哺乳類。61 (2): 376–377. ドイ:10.2307/1380075。JSTOR 1380075 .
^ Jordheim、S(1980)。「イーグルコヨーテ事件」 . ブルージェイ。38 : 47–48. ドイ: 10.29173/bluejay4577 .
^ ユング、TS。エバラット、KT; Andresen-Everatt、LM (2009)。「カナダ北西部のイヌワシ (Aquila chrysaetos) によるコヨーテ (Canis latrans) の盗寄生」. 北西部のナチュラリスト。90 (1): 53–55. ドイ: 10.1898/1051-1733-90.1.53。S2CID 86356027 .
^ マサチューセッツ州マッコロー。トッド、CS。オーウェンRBジュニア(1994)。「メイン州で越冬するハクトウワシのための補助給餌プログラム」. 野生生物協会の会報。22 (2): 147–154. ドイ:10.1016/0006-3207(96)83230-7 . JSTOR 3783240 .
^ 軍曹、アラン・B。アレン、スティーブン H. (1989)。「コヨーテとアカギツネの相互作用を観察」. ジャーナル オブ マンマロジー. 70 (3): 631–633. ドイ:10.2307/1381437。ISSN 1545-1542。JSTOR 1381437 . OCLC 39098574。
^ フェドリアーニ、JM。フラー、TK。サウバジョット、RM; ヨーク、EC (2000)。「同所性肉食動物 3 種の競争とギルド内捕食」 (PDF) . オエコロジー。125 (2): 258–270. ビブコード: 2000Oecol.125..258F . ドイ: 10.1007/s004420000448 . hdl : 10261/54628。ISSN 1432-1939 . OCLC 76327396。PMID 24595837。S2CID 24289407 . 2011 年 10 月 6 日にオリジナル (PDF)からアーカイブされました。
^ ジャワ州リトバイティス。DJ ハリソン (1989)。「コヨーテの個体数が増加する期間中のボブキャットとコヨーテのニッチな関係」. カナダ動物学ジャーナル。67 (5): 1180–1188。ドイ:10.1139/z89-170。
^ Bunnell、Kevin D.; フリンダース、ジェラン T.; ウルフ、マイケル L. (2006)。「山間西部のオオヤマネコの保護に対するコヨーテとスノーモービルの潜在的な影響」. 野生生物協会の会報。34 (3): 828–838. doi : 10.2193/0091-7648(2006)342.0.CO;2。ISSN 1938-5463。JSTOR 3784713 . OCLC 60353682。S2CID 85901240 . ^
PS ギプソン; カムラー、JF (2002)。「コヨーテに殺されたボブキャット」。南西部のナチュラリスト。47 (3): 511–513. ドイ:10.2307/3672519。ISSN 0038-4909 . JSTOR 3672519 . OCLC 525604174。
^ アンダーソン、EM (1986). コロラド州南東部における資源利用に関連するボブキャットの行動生態学。論文、コロラド州立大学、フォートコリンズ、米国。
^ ジャクソン、DH (1986). コロラド州中東部のボブキャットの生態。論文、コロラド州立大学、フォートコリンズ、米国。
^ Toweill, DE (1986). 針葉樹林におけるボブキャットとコヨーテによる資源分配。論文、オレゴン州立大学、コーバリス、米国
^ ニック、ST (1990)。「アイダホ州南東部における搾取と獲物の減少に関連するボブキャットの生態」. 野生生物のモノグラフ。108 (108): 1–42. JSTOR 3830671 .
^ リップル、WJ。Wirsing、AJ。ベシュタ、RL。ブスカーク、SW (2011)。「オオカミを元に戻すことはオオヤマネコの回復を助けることができますか?」 (PDF) . 野生生物協会の会報。35 (4): 514–518. ドイ: 10.1002/wsb.59 .
^ オドノヒュー、M.; ホーファー、EJ。ドイル、FI(1995)。「捕食者対捕食者」。自然史。104 : 6–9.
^ ロックウッド、ラリー L. (2015)。集団生態学の紹介。イギリス、チチェスター:ジョン・ワイリーと息子たち。p。273.ISBN _ 978-1-118-94755-5. OCLC 899267730。
^ ブッシュカーク、SW; ルッジェーロ、LF; クレブス、CJ (2000)。「生息地の断片化と種間競争: オオヤマネコの保全への影響」 (PDF) . ルッジェーロ、LF。オーブリー、KB。バスカーク、サウスウェスト; ケーラー、GM。クレブス、CJ。カンザス州マッケルビー。スクワイアーズ、JR (eds.)。米国におけるオオヤマネコの生態と保全。デンバー: コロラド大学出版局。pp.91–92。
^ ホーディ、ジェームズ・W; ケイズ、ローランド(2018)。「北米および中央アメリカにおけるコヨーテ (Canis latrans) の拡大のマッピング」 . ZooKeys (759): 81–97。ドイ: 10.3897/zookeys.759.15149 . PMC 5974007 . PMID 29861647。
^ 「コヨーテは北アメリカを征服しました。今、彼らは南に向かっています」 . ニューヨークタイムズ。2018 年 5 月 24 日。ISSN 0362-4331。2018 年 5 月 24 日にオリジナルからアーカイブされました。2018年5 月 25 日閲覧。
^ Hidalgo-Mihart、MG (2004)。「メキシコと中央アメリカにおけるコヨーテ ( Canis latrans )の歴史的および現在の分布」 . 生物地理学のジャーナル。31 (12): 2025–2038. ドイ: 10.1111/j.1365-2699.2004.01163.x . S2CID 56048806 . 2017 年 5 月 25 日にオリジナルからアーカイブされました。2015年1 月 25 日閲覧。
^ デラロサ、CL; Nocke、CC(2010)。「肉食動物の進化:中央アメリカと南北の大移動」 . 中央アメリカの肉食動物ガイド: 自然史、生態学、保全。テキサス大学プレス。ISBN 978-0-292-78951-7.
^ ヘッペンハイマー、エリザベス。コシオ、ダニエラ S.; Brzeski、Kristin E.; コーディル、ダニー。ヴァン・ホワイ、カイル。チェンバレン、マイケル J.; ヒントン、ジョセフ W.; ボンホールド、ブリジット(2017)。「最近拡大したコヨーテ (Canis latrans) の個体群では、人口学的歴史が遺伝的多様性の空間パターンに影響を与える」 . 遺伝。120 (3): 183–195. ドイ: 10.1038/s41437-017-0014-5 . PMC 5836586 . PMID 29269931。
^ ヤング & ジャクソン 1978 , pp. 107–114
^ Gier、HT。クラッケンバーグ、SM; マーラー、RJ (1978)。「コヨーテの寄生虫と病気」. In Bekoff, M. (ed.)。コヨーテ: 生物学、行動、および管理。ニューヨーク:アカデミックプレス。pp.37–71。ISBN 978-1-930665-42-2. OCLC 52626838。
^ チャイ、JY; ダーウィン、マレル K.; AJ リンベリー (2005)。「魚介類寄生性人畜共通感染症: 状況と問題」. 寄生虫学のための国際ジャーナル。35 (11–12): 1233–1254。ドイ: 10.1016/j.ijpara.2005.07.013 . PMID 16143336。
^ ワッツ、LS (2006)。アメリカの民間伝承の百科事典。インフォベース出版。pp.93–94。ISBN 978-1-4381-2979-2. OCLC 465438817。
^ ハリス、M. (1979)。文化的唯物論:文化の科学のための闘争。ニューヨーク:AltaMira Press。pp.200–1。ISBN 978-0-7591-0135-7. OCLC 47100657。
^ ギレスピー、アンガス K.; Mechling、ジェイ(1987)。シンボルとストーリーにおけるアメリカの野生生物。テネシー大学出版局。pp.225–230 。_ ISBN 978-0-87049-522-9. OCLC 14165533。
^ ウェイ、JG(2012)。「オオカミが好きでコヨーテが嫌いなのか? イヌ科愛好家の難問」 (PDF) . インターナショナルウルフ。22 (4): 8–11. 2012 年 12 月 24 日にオリジナル (PDF)からアーカイブされました。
^ ペンシルベニア州リンチ。ロバーツ、J. (2010)。ネイティブ アメリカンの神話 A から Z . インフォベース出版。p。27.ISBN _ 978-1-4381-3311-9. OCLC 720592939。
^ Schwartz, M. (1998). 初期アメリカ大陸における犬の歴史。イェール大学出版局。pp.146–149。
ISBN 978-0-300-07519-9 .
^ ミラー、メイン。タウベ、カザフスタン (1993)。古代メキシコとマヤの神々とシンボル: メソアメリカ宗教の図解辞書. テムズとハドソン。p。 92 . ISBN 978-0-500-05068-2. OCLC 27667317。
^ オリビエ、G. (2003)。アステカの神の嘲笑と変容:テスカトリポカ、「喫煙鏡の主」 . コロラド大学出版局。p。 32 . ISBN 978-0-87081-745-8. OCLC 52334747。
^ ケリー、DH (1955)。「ケツァルコアトルと彼のコヨーテの起源」. エル メキシコ アンティグオ。8 : 397–416. ^
「コヨーテの攻撃: 増加する郊外の問題」 (PDF) . 2004 年 3 月。2007 年9 月 26 日のオリジナル (PDF)からのアーカイブ。2007年8 月 19 日閲覧。
^ アタック イン ザ ワイルド: コヨーテ ミステリー(ドキュメンタリー)。ナショナル ジオグラフィック チャンネル。2009 年 10 月 27 日。2016年11 月 5 日閲覧。
^ Dell’Amore、Christine 。「シティスリンカーズ」。スミソニアン。2012年6 月 14 日閲覧。
^ ベイカー、レックス O. (2007)。「南カリフォルニアで人間やペットへの攻撃を防止または軽減するために実施された成功した都市コヨーテ管理プログラムのレビュー」 . 第 12 回野生生物被害管理会議の議事録。pp.382–392。
^ ケイズ、R。カーティス、A。Kirchman、JJ(2009)。「オオカミとの交配による北東コヨーテの急速な適応進化」 . 生物学の手紙。6 (1): 89–93. ドイ: 10.1098/rsbl.2009.0575 . PMC 2817252 . PMID 19776058 . ^
「コヨーテの捕食 – 説明」 . 2007 年 8 月 6 日にオリジナルからアーカイブされました。2007年8 月 19 日閲覧。
^ 羊とヤギの死の損失 (PDF) (レポート)。全国農業統計局。2005 年 5 月 6 日。2020年11月27日閲覧。
^ 羊とヤギの死の損失 (PDF) (レポート)。全国農業統計局。2010 年 5 月 27 日。2020年11月27日閲覧。
^ 「米国におけるヒツジと子羊の捕食者と非捕食者の死の損失、2015年」 (PDF) . 米国農務省。2015 . 2016年7 月 1 日閲覧。
^ 「羊と子羊の在庫」 . 米国農務省。2010年2月1日閲覧。
^ 「狡猾なコヨーテを操る? まだ簡単な答えはない」 . NBCニュース。2009 年 12 月 7 日。2013年9 月 14 日閲覧。
^ コリンジ、マーク。ティム、RM; マドン、MB (2008)。「アイダホ州の個々のオオカミ、ツキノワグマ、ピューマ、コヨーテによる家畜の捕食の相対的リスク」 . 脊椎動物害虫会議の議事録: 129–133。
^ ユタ州のプレデター コントロール プログラムの概要、2016 年 7 月 1 日から 2017 年 6 月 30 日までのプログラム活動とデータ (PDF) (レポート)。野生生物資源のユタ部門。2017年。
^ 「家畜保護犬」 (PDF) . 野生生物サービス。2010 年 10 月。2016年 9 月 9 日にオリジナル (PDF)からアーカイブされました。2016年7 月 3 日閲覧。
^ 「家畜番犬ファクトシート」 . 動物および植物の健康検査サービス米国農務省。2012 年 3 月 24 日にオリジナルからアーカイブされました。2012年4 月 3 日閲覧。
^ グランディン、テンプル (2015 年 2 月 26 日)。「専門家は、放牧を正しく行うと、環境と野生生物の生息地が改善されると言っています」 . ビーフマガジン。2017年12 月 30 日閲覧。
^ 「飼育された牛、飼育された未経産牛、フィーダーの子牛」 . Rhino’s Beef & Farm Raised Aussies . 2016 年 10 月 4 日。2017年12 月 30 日閲覧。
^ 「オオカミの略奪への牧場主のガイド」 . モンタナ州立大学。2006. 2013 年 4 月 9 日時点のオリジナルからのアーカイブ。2016年7 月 1 日閲覧。
^ ローリンズ、デール。「コヨーテに対処する: 家畜の損失を最小限に抑えるための代替管理」 (PDF) . テキサス農業普及サービス。pp. 4–7 。2016年11 月 5 日閲覧。
^ Hardesty、グレッグ (2005 年 5 月 5 日)。「コヨーテにとってペットは獲物」 . グレッグ・ハーデスティ。オレンジカウンティレジスター. 2007 年 7 月 15 日にオリジナルからアーカイブされました。
^ 「ウェイマスでコヨーテが攻撃し、犬を殺す」 . WHDH-TV – ニューイングランド ニュース。2007 年 5 月 14 日
^ マクール、ジュリエット(2010)。「コヨーテ対グレイハウンド:バトルラインが描かれている」 . ニューヨークタイムズ。2022 年 1 月 1 日にオリジナルからアーカイブされました。2016年7 月 3 日閲覧。
^ 「コヨーテ・ネーション:コヨーテ狩りの短期集中コース」 . アウトドアライフ。2019 年 1 月 23 日。2019年12月22日閲覧。
^ 「コヨーテ・ネーション:コヨーテ狩りの短期集中コース」 . アウトドアライフ。2019 年 1 月 23 日。2020年5月7日閲覧。
^ 「コヨーテの足跡の見分け方」 . ワイルドライフ ランド トラスト。2020年5月10日閲覧。
^ Young & Jackson 1978 , pp. 115–116 ^ セトン 1909 年、p. 816 ^ ヤング & ジャクソン 1978 , pp. 119–21
^ ヤング & ジャクソン 1978 , pp. 64–9
^ シュルツ、JW(1962)。Blackfeet and Buffalo: Memories of Life Among the Indians . オクラホマ大学出版局。pp.141–3。ISBN 978-0-8061-1700-3. OCLC 248716 .
^ Etter, J. (1998 年 2 月 15 日)。「コヨーテはダーラムマンの最高のバードドッグとして溶け込む」 . オクラホマン。2016年7 月 1 日閲覧。
^ エドワーズ、マーク (2007 年 4 月 1 日)。「善人、勇者、華麗なる者」 . タイムズ。ロンドン。2011 年 5 月 17 日にオリジナルからアーカイブされました。2007年5 月 26 日閲覧。
一般的な情報源
Cartaino、キャロル (2011)。コヨーテに関する神話と真実: アメリカで最も誤解されている捕食者について知っておくべきこと. howyouwant.comを読んでISBN 978-1-4587-2668-1. OCLC 876517032。
Fox、MW(1978)。犬:その家畜化と行動。ガーランド STPM プレス。ISBN 978-0-8240-9858-2. OCLC 3223381 .
ジョンストン、CS(1938)。「チタ キャニオンの脊椎動物型産地と祖先コヨーテの記述に関する予備報告」. アメリカ科学ジャーナル。5. 35 (209): 383–390. ビブコード: 1938AmJS…35..383J . ドイ: 10.2475/ajs.s5-35.209.383 .
Nowak、RM (1979)。「最近の人口の歴史と統計分析」。In Wiley, EO (ed.)。北米第四紀カニス. 巻。6. カンザス州ローレンス: カンザス大学印刷サービス。ISBN 0-89338-007-5.
Nowak、RM (2003)。「オオカミの進化と分類学」. Mechでは、L. David。ボイタニ、ルイージ(編)。オオカミ: 行動、生態学および保全。シカゴ大学出版局。pp.239–258。ISBN 978-0-226-51696-7.
セトン、ET(1909)。北方動物の生活史:マニトバの哺乳類の説明。ニューヨーク:スクリブナー。
テッドフォード、リチャード・H。ワン、シャオミン。テイラー、ベリル E. (2009)。「北米化石イヌ科(食肉目:イヌ科)の系統発生学」 (PDF) . アメリカ自然史博物館紀要。325 : 1–218. ドイ:10.1206/574.1。hdl : 2246/5999。S2CID 83594819 .
ワン、シャオミン。テッドフォード、リチャード H. (2008)。犬: その化石の親戚と進化の歴史. ニューヨーク:コロンビア大学出版局。ISBN 978-0-231-13528-3. OCLC 185095648。
ヤング、SP。ジャクソン、HHT (1978)。賢いコヨーテ。ネブラスカ大学出版局。ISBN 978-0-8032-5893-8. OCLC 3294630。
参考文献編集
書籍
ディクソン、JS(1920)。カリフォルニアでのコヨーテの管理。カリフォルニア州バークレー : 農業試験場
フローレス、D. (2016)。コヨーテアメリカ:自然史と超自然史。ベーシックブック。
ISBN 978-0-465-05299-8
アーカンソー州ハーディング (1909)。オオカミとコヨーテの捕獲; 経験豊富な「オオカミ」がこれらの動物を狩り、罠にかけるための最も成功した方法を提供する最新のオオカミハンターのガイドは、彼らの習慣も詳細に提供します. コロンバス、オハイオ、AR ハーディング パブ。共同。
クルテン、B (1974)。「コヨーテに似た犬(イヌ科、哺乳類)の歴史」. Acta Zoologica Fennica . 140 : 1–38.
Leydet、フランソワ (1988)。コヨーテ:西部の挑戦的なソングドッグ。オクラホマ大学出版局。ISBN 978-0-8061-2123-9. OCLC 17106424。
モリー、ポール(2004)。シカゴ大都市圏におけるコヨーテ Canis latrans の景観利用と食事(論文)。ユタ州立大学。
Murie, A. (1940)。イエローストーンのコヨーテの生態。ワシントンDC:USGPO
パーカー、ジェリー。(1995)。「Eastern Coyote: Story of Its Success」、Nimbus Publishing、ハリファックス、ノバスコシア、カナダ。
ヴァン・ナイス、フランク(2015)。被害者と犠牲者:アメリカ西部における捕食者の管理。カンザス州ローレンス: カンザス大学出版局。
ワーグナー、MM(1920年頃)。飼いならされたコヨーテの自伝。サンフランシスコ、ハーワーグナーパブ。共同。
ビデオ
シェリー、プリヤ 。コヨーテと暮らす(18分)。イオン。
オーディオブック
オルソン、ジャック 。最後のコヨーテ(8 時間)。ゲイリー・マクファーデンによるナレーション。原書はSlaughter the Animals, Poison the Earth , Simon & Schuster, 1971年10月11日。ASIN B00WGUA5HK。
外部リンク![]()
ウィキクォートにコヨーテ
に関する引用句集が
![]()
ウィキスピーシーズにCanis latransに関する情報が
![]()
コヨーテを調べて![]()
・コモンズには、コヨーテに関連するカテゴリが
「コヨーテ」 。ブリタニカ百科事典。巻。7 (第 11 版)。1911年。
「カニス・ラトランス」。統合分類情報システム。2006年3 月 23 日閲覧。
アリゾナ ゲーム & フィッシュ部門、「コヨーテと暮らす」
西洋コヨーテ、オオカミ、コヨーテ DNA バンク @ トレント大学
生物多様性遺産図書館でCanis latransの発生を表示します。”