Plains_coyote
その他の用法については「コヨーテ 」をご覧
コヨーテ( Canis latrans ) は、北アメリカ原産のイヌ科の一種です。近縁種であるオオカミよりも小さく、近縁種であるヒガシオオカミやアカオオカミよりもわずかに小さいです。ユーラシアにおけるキンイロジャッカルと同じ生態的地位の多くを占めています。コヨーテはより大型で捕食性が高く、かつては行動生態学者によってアメリカンジャッカルと呼ばれていました。この種の歴史的な名前には他に、プレーリーオオカミやブラッシュオオカミなどが
コヨーテ
時間範囲:中期更新世– 現在 (0.74–0.85 Ma )
マウンテンコヨーテ ( C. l. lestes )
保存状況
最小懸念 ( IUCN 3.1 )
科学的分類
王国:
動物界
門:
脊索動物
クラス: 哺乳類 注文: 食肉目 家族: イヌ科
属: カニス 種族:
C.ラトランス
二項名
カニス・ラトランス
セイ、1823年
Canis latransの最新シリーズ リスト
カニス・アンダーソーニ ・メリアム、1910
Canis caneloensisスキナー、1942 年
イヌ クレプティカスエリオット、1903 年
カニス・エストール・ メリアム、1897
Canis フラストラー ウッドハウス、1851 年
カニス・ゴールドマニ ・メリアム、1904
Canis hondurensis ゴールドマン、1936
Canis impavidus アレン、1903 年
Canis irvingtonensis Savage、1951 年
Canis jamesiタウンゼント、1912 年
イヌ レステス メリアム、1897
Canis mearnsi Merriam、1897
イヌ ミクロドン メリアム、1897
Canis nebrascensis Merriam、1898
Canis ochropus Eschsholtz、1829
Canis orcutti Merriam、1910
Canis pallidus メリアム、1897
カニス半島 メリアム、1897
Canis riviveronis ヘイ、1917
Canis vigilis メリアム、1897
Lyciscus cagottisハミルトン=スミス、1839 年
コヨーテは、北米全域に広く分布し、豊富に生息しているため、国際自然保護連合によって最も懸念されていない動物としてリストされています。この種は多用途であり、人間によって改変された環境に適応して拡大することができます。都市部のコヨーテは多くの都市でよく見られます。コヨーテは2013年にパナマ東部(生息地からパナマ運河を渡ったところ)で初めて目撃された。
コヨーテには 19 の亜種が認められています。平均的なオスの体重は 8 ~ 20 kg (18 ~ 44 ポンド)、平均的なメスの体重は 7 ~ 18 kg (15 ~ 40 ポンド) です。毛皮の色は主にライトグレーと赤、または黒と白が散りばめられたフルボッシュですが、地理によって多少異なります。社会組織において非常に柔軟性があり、家族単位、または血縁関係のない個人の緩やかな群れで生活します。主に肉食で、主にシカ、ウサギ、ノウサギ、げっ歯類、鳥、爬虫類、両生類、魚、無脊椎動物を主食としていますが、果物や野菜も食べることも特徴的な鳴き声は孤独な個体が発する遠吠えです。コヨーテの最大の脅威は人間であり、クーガーとハイイロオオカミがそれに続きます。それにもかかわらず、コヨーテはハイイロオオカミ、ヒガシオオカミ、またはレッドオオカミと交尾し、「コイウルフ」のハイブリッドを生み出すことが北米の北東部地域では、イースタンコヨーテ(より大型の亜種ですが、それでもオオカミより小さい)は、さまざまな種類のオオカミとの歴史的および最近のさまざまな交配の結果です。遺伝子研究によると、北米のオオカミのほとんどは、ある程度のコヨーテの DNA を持っていることがわかっています。
コヨーテは、主にアリドアアメリカのネイティブ アメリカンの民間伝承の著名なキャラクターであり、通常は実際のコヨーテまたは人間の姿を交互にとるトリックスターとして描かれます。他のトリックスターの人物と同様に、コヨーテは欺瞞とユーモアを使って社会的慣習に反逆します。この動物はメソアメリカの宇宙論において軍事力の象徴として特に尊敬されていました。ヨーロッパがアメリカ大陸を植民地化した後、英米文化では臆病で信頼できない動物とみなされていました。世間のイメージが向上したオオカミとは異なり、コヨーテに対する態度は依然として否定的なものが多い。
コンテンツ
1 説明
2 分類と進化
2.1 歴史 2.2 ネーミングと語源 2.3 進化
2.3.1 化石記録
2.3.2 DNAの証拠
2.4 亜種 2.5 ハイブリダイゼーション
3 行動
3.1 社会的および生殖行動 3.2 縄張りと避難行動 3.3 狩猟と採食行動 3.4 コミュニケーション
3.4.1 ボディランゲージ
3.4.2 発声
4 エコロジー
4.1 生息地 4.2 ダイエット 4.3 敵と競争相手
5 範囲
6 病気と寄生虫
7 人間との関係
7.1 民間伝承や神話では 7.2 人間への攻撃 7.3 家畜やペットの捕食 7.4 狩猟 7.5 毛皮の用途 7.6 飼い慣らしやすさ 7.7 人気メディアで
8 解説
9 引用
10 一般的な情報源と引用された情報源
11 参考文献
11.1 本 11.2 ビデオ 11.3 オーディオブック
12 外部リンク
説明

マウンテン コヨーテ ( C. l. lestes ) の頭
の拡大図
コヨーテのオスの体重は平均 8 ~ 20 kg (18 ~ 44 ポンド)、メスの体重は平均 7 ~ 18 kg (15 ~ 40 ポンド) ですが、サイズは地理的に異なります。平均体重 18 kg (40 ポンド) の北部亜種は、平均 11.5 kg (25 ポンド) のメキシコ南部亜種よりも大きくなる傾向が全長は平均して 1.0 ~ 1.35 m (3 フィート 3 インチ~4 フィート 5 インチ) です。尾の長さは 40 cm (16 インチ) で、メスの方が体長も体高も短いです。記録上最大のコヨーテは、1937年11月19日にワイオミング州アフトン近郊で殺されたオスで、鼻から尾までの長さは1.5メートル(4フィート11インチ)、体重は34キロ(75ポンド)であった。臭腺は尾の付け根の上側にあり、青黒い色をしています。
コヨーテの毛皮の色と質感は地理的に多少異なります。毛の主な色は明るい灰色と赤またはフルーヴスで、体の周りには黒と白が点在しています。高地に生息するコヨーテは、砂漠に生息するコヨーテの濃い灰色または白っぽい灰色に比べて、黒と灰色の色合いが多い傾向がコヨーテの毛皮は、短くて柔らかい下毛と、長くて粗いガード毛で構成されています。北方の亜種の毛皮は南方の亜種よりも長く密度が高く、メキシコと中央アメリカの一部の亜種の毛皮はほとんど茶色(剛毛)です。一般に、成体のコヨーテ(コイウルフのハイブリッドを含む)はクロテンの毛色、新生児の濃い毛色、活動的な尾上腺を備えたふさふさした尾、および白い顔のマスクを持っています。 コヨーテでは白皮症は非常にまれです。1938年3月から1945年6月までの間に連邦政府と狩猟協同組合によって殺された合計75万頭のコヨーテのうち、アルビノはわずか2頭であった。
コヨーテは通常、ハイイロオオカミよりも小さいですが、耳が長く、脳ケースが比較的大きく、フレーム、顔、銃口が薄いのが特徴です。臭腺はハイイロオオカミより小さいですが、同じ色です。毛皮の色のバリエーションはオオカミに比べてはるかに少ない。コヨーテは走ったり歩いたりするときに、オオカミのように尻尾を水平にではなく、下に下げます。
コヨーテの足跡は、より細長く、丸みが少ない形状によって犬の足跡と区別できます。 イヌとは異なり、コヨーテの上部犬歯はオトガイ孔を越えて伸びている。
分類と進化編集
歴史

コヨーテの
トルテカ族の絵文字
ヨーロッパ人がアメリカ大陸を植民地化した当時、コヨーテは主に大陸の西半分の開けた平原と乾燥地帯に限られていました。コロンブス以降の初期の歴史記録では、作家がコヨーテについて記述しているのか、それともオオカミについて記述しているのかを判断するのは困難な場合が多い。イリノイ州カスカスキアで地元の司祭が書いた1750年の記録には、そこで遭遇した「オオカミ」はヨーロッパのオオカミよりも小さく、勇敢さがなかったと記されている。エドワーズ郡の1800年代初頭の別の記述では、オオカミが夜に遠吠えしていると述べられているが、これらはコヨーテである可能性が高い。この種はルイスとクラークの遠征(1804年から1806年)中に数回遭遇したが、ミズーリ州上流のヨーロッパの貿易業者にはすでによく知られていた。1805 年 5 月 5 日にモンタナ州北東部で書いたメリウェザー・ルイスは、コヨーテを次のような言葉で描写しました。
草原の小さなオオカミまたは穴を掘る犬は、ほぼ例外なく、開けた平原の住民です。彼らは通常、10 人か 12 人の群れを作り、時にはそれ以上で行動し、獲物が頻繁に訪れる峠や場所の近くに巣を作ります。鹿やヤギを一人で捕まえることができないため、単独で見つかることはほとんどなく、群れで狩りをします。彼らは頻繁に巣穴の近くで獲物を観察して捕まえます。これらの巣穴で彼らは子を育てますが、追いかけられたときも彼らに頼ります。人が近づくとよく吠えますが、その声はまさに小型犬のものです。彼らはキツネとイヌの中間的な大きさで、非常に活発な船団で繊細に形成されています。耳は大きく直立し、頭は長くてキツネのように尖っていました。長い話です…髪と毛皮もキツネに似ていますが、はるかに粗くて劣っています。それらは淡い赤褐色をしています。深海緑色の小さくて鋭い瞳。彼らのは普通のオオカミのものや大西洋諸国に一般的なものよりかなり長いですが、そのどれもこの地区では見つけることができませんし、プラット川の上では私は信じ
コヨーテは、 1819年9月に博物学者の トーマス・セイによって、政府主催のスティーブン・ロング少佐との遠征中に、プラット河口からミズーリ川を24キロ上流したルイス・アンド・クラーク評議会ブラフスの敷地で、初めて科学的に記載された。。彼はルイスとクラークのジャーナルの初版を手元に持っており、そこには 1805 年 5 月 5 日付のルイスの観察をビドルが編集したバージョンが含まれていました。彼の記述は 1823 年に出版されました。セイは「プレーリーオオカミ」の違いを文書化した最初の人物でした。コヨーテ)、そして彼の日記の次のページには、 Canis nubilus ( Great Plains wolf )と名付けたオオカミが描かれています。 セイはコヨーテについて次のように説明した。
イヌ ラトランス。銀色または灰色、上は黒、鈍いフルーヴォまたはシナモンと変化します。髪の根元は薄暗く、長さの中央はくすんだシナモン色で、先端は灰色または黒で、脊椎線に沿って長くなります。耳は直立し、先端は丸く、後ろはシナモン色、髪の根元は黒く太く、内側には白髪が並んでいます。まぶたは黒で縁取られ、上まつげは下と上の先端が黒くなっています。補助蓋は前が黒茶色で縁取られ、後ろが黒茶色で縁取られています。アイリスイエロー。瞳孔は黒青。涙嚢に黒褐色の斑点がある。鼻のシナモン、鼻に灰色がかった色がかった色。唇は白く、黒で縁取られ、3連の黒いセタ。耳の間の頭は灰色とくすんだシナモン色が混ざり、根元の毛はくすんで太い。側面は背中よりも青白く、脚の上は時代遅れの黒で覆われています。脚の外側はシナモン色で、後部の髪がよりはっきりしています。手首近くの前部の拡張された黒い短縮線。尾はふさふさしており、紡錘形で、真っ直ぐで、灰色とシナモン色で変化しており、上部の基部近くに斑点があり、先端は黒色です。尾の胴の先端は、脚を伸ばすと尾根の先端に達します。下は白く、真っ白で、先端に向かって尾はシナモン色で、先端は黒です。後ろ足は4本指、前足は5本指。
ネーミングと語源
この種について書かれた最古の言及は、博物学者 フランシスコ・エルナンデスの 『植物と動物』(1651年)にあり、そこでは「スペインのキツネ」または「ジャッカル」として説明されています。「コヨーテ」という言葉 (ナワトル語名コヨートルの 発音(ヘルプ・情報)をスペイン語で借用したもの) が最初に出版されたのは、1780 年の歴史家フランシスコ・ハビエル・クラビジェロの『ヒストリア・デ・メキシコ』にある。 英語で使用されたのはウィリアム・ブロックの6か月間のメキシコ滞在と旅行(1824年)であり、そこではcayjotteやcocyotieなど様々に転写されている。この単語の綴りは 1880 年代までに「coyote」として標準化されました。
![]() コヨーテの別の英語名には、「プレーリーウルフ」、「ブラッシュウルフ」、「ケースドウルフ」 、 「リトルウルフ」、「アメリカンジャッカル」などがある。その二項名 Canis latrans は「吠える犬」を意味し、彼らが発する多くの鳴き声にちなんでいます。 Canis latrans の地元および先住民の名前
コヨーテの別の英語名には、「プレーリーウルフ」、「ブラッシュウルフ」、「ケースドウルフ」 、 「リトルウルフ」、「アメリカンジャッカル」などがある。その二項名 Canis latrans は「吠える犬」を意味し、彼らが発する多くの鳴き声にちなんでいます。 Canis latrans の地元および先住民の名前
言語グループまたは言語領域
先住民の名前
アリカラ ストシリア・プカチュ
カナダフランス語 コヨーテ
チヌーク
イタリパス
チペウィアン Nu-ní-yĕ=̑ts!ế-lĕ
ココパ シュパシュパ
クリー北部クリー平原クリー ᒣᐢᒐᒑᑲᓂᐢ (ミスカッカカニス) ᒣᐢᒐᒑᑲᓂᐢ (メスカカニス)
クリーク Yv・hu・ce (古風) Yv・hv・la・nu・ce (現代)
ダコタ マイカミカクシカ
コチ シンチレップ
ヒダツァ モツァ
ホピ族 イイサウ イサウ
カルク ピネフィッチ
クラマス コ・ハ・ア
マンダン シェケ
マヤ語 ペキキャッシュ
ネズ・パース アイスエイ・イェ
ナワトル島 コヨートル
ナバホ族 マイイ
オガララ・スー ミーヤスレイチャラ
オジブワ語 ᒣᔅᑕᒐᐦᑲᓀᔅ メスタチャガンエス
オマハ ミカシ
オーセージ ó ͘ ショムカシ
ポーニー キキリキ
ピウテ エジャア
スペイン語 コヨーテペロ・デ・モンテ
やかま テリパ
ティンビシャ イサ(プ)イサパイプイサパイプ
ウィントゥ チャラワセデト
ヤンクトン・スー ソントケチャ
ユロック セゲプ
進化
数百万年単位のタイミングによるオオカミに似たイヌ科動物の系統樹
イヌ科 3.5 Ma3.0 2.5 2.0 0.96 0.6 0.38
飼い犬
![]()
ハイイロオオカミ
![]()
コヨーテ
![]()
アフリカオオカミ
![]()
キンイロジャッカル
![]()
エチオピアオオカミ
![]()
ドール
![]()
アフリカの野生の犬
![]()
2.6 横縞ジャッカル
![]()
セグロジャッカル
![]()
化石記録
参照:オオカミの進化 § 化石記録
肉食動物の進化の第一人者であるシャオミン・ワンとリチャード・H・テッドフォードは、イヌ属はコヨーテに似たユーキオン・ダビシの子孫であり、その遺骸は600万年前の中新世に初めて出現したと提案した(Mya)米国南西部とメキシコで。鮮新世(500 万年)までに、より大型のCanis lepophagus が同じ地域に出現し、更新世初期(100 万年)までにはC. latrans (コヨーテ) が存在していました。彼らは、 Eucyon davisiからC. lepophagus、そしてコヨーテへの進化は直線進化であると提案した。
C. latransとC. aureusはC. edwardiiと密接に関連しており、C. edwardiiはブランカン中部(鮮新世後期) からアーヴィントン紀末 (更新世後期)にかけて最も早く出現した種であり、 C. latransと区別できないコヨーテの死骸は同時代のものでした。北米のC. edwardiiと。ジョンストンは、C. lepophagus は現代のコヨーテよりも細い頭蓋骨と骨格を持っていると説明しています。ロナルド・ノワクは、初期の個体群は小型のコヨーテに似た小さく繊細な狭い比率の頭蓋骨を持ち、C. latransの祖先であると思われることを発見した。
C. lepophagus は現代のコヨーテと体重は似ていましたが、四肢の骨が短く、あまり大雑把なライフスタイルを示していました。コヨーテはハイイロオオカミよりも原始的なイヌ科の動物であり、その比較的小さいサイズと比較的狭い頭蓋骨と顎が示すように、オオカミが得意とする大きな獲物を掴むのに必要な把握力が欠けています。これは、コヨーテの矢状稜が低いか完全に平坦であることからもさらに裏付けられており、オオカミよりも噛みつきが弱いことを示しています。コヨーテはオオカミのような特殊な肉食動物ではなく、奥歯の咀嚼面が大きいことからわかるように、この種が植物質に相対的に依存していることを反映しています。これらの点で、コヨーテはオオカミよりもキツネに似たこの属の祖先に似ています。
現生コヨーテの範囲に含まれる最古の化石は、ウェストバージニア州ハミルトン洞窟で074~085万年前に発見された。カリフォルニア州アービントンでは0.73Ma。コロラド州ポーキュパイン洞窟とペンシルバニア州カンバーランド洞窟では0.35〜0.48Ma。現代のコヨーテは、第四紀の絶滅事件から 1,000 年後に出現しました。現代の完新世のコヨーテと比較して、更新世のコヨーテ( C. l. orcutti ) はより大きく、より頑丈であり、より大きな競争者や獲物に反応したと考えられます。更新世のコヨーテは、その歯が肉を切るのに適応しており、植物を処理するのに適した研磨面が少ないため、その子孫よりも肉食に特化した動物であった可能性が高い。彼らのサイズの縮小は、彼らの大きな獲物が絶滅した第四紀の絶滅イベントから1,000年以内に起こりました。さらに、更新世のコヨーテは、ダイアオオカミ( Aenocyon dirus )の絶滅後に空いた大物狩猟のニッチを利用することができませんでした。なぜなら、このニッチはハイイロオオカミによって急速に埋められ、おそらくハイイロオオカミが大型コヨーテを積極的に殺したと思われます。現代のしなやかな形態を好む自然選択。
DNAの証拠
![]()
更新世のコヨーテ( C. l. orcutti )
の骨格
1993年、ある研究では、北米のオオカミはユーラシアのオオカミよりもコヨーテに似た頭蓋骨の特徴を示していると提案されました。 2010年の研究では、コヨーテは中国チベットオオカミ、飼い犬、モンゴルオオカミ、ユーラシアオオカミを含むクレードの基本的なメンバーであり、中国チベットオオカミはオオカミと飼い犬から早期に分岐したことが判明した。
2016年、全ゲノムDNA研究により、仮定に基づいて、北米のすべてのオオカミとコヨーテは約5万1000年前に共通の祖先から分岐したと提唱された。 しかし、提案されているオオカミとコヨーテの分岐時期は、100万年前の地層でコヨーテに似た標本が発見されたことと矛盾している。また、この研究では、北米のすべてのオオカミはかなりの量のコヨーテの祖先を持ち、すべてのコヨーテはある程度のオオカミの祖先を持っており、アカオオカミとヒガシオオカミには異なる割合でハイイロオオカミとコヨーテの祖先が高度に混合されていることが示された。
オオカミやイヌに関する遺伝研究では、入手可能な唯一の参照ゲノムであるボクサー犬のゲノムに基づいて系統関係が推測されています。2017 年、将来の研究に役立てるため、オオカミCanis lupus lupusの最初の参照ゲノムがマッピングされました。 2018年、ある研究では、北アメリカのオオカミ、オオカミに似たイヌ科動物、コヨーテのゲノム構造とその混合を、オオカミの参照ゲノムに対して核ゲノム配列の最大のデータセットをマッピングした全範囲にわたる標本を用いて調べた。
この研究は、北米のハイイロオオカミとオオカミに似たイヌ科動物がハイイロオオカミとコヨーテの複雑な混合の結果であるというこれまでの研究結果を裏付けるものである。グリーンランドのホッキョクオオカミとメキシコのコヨーテは最も純粋な標本の代表でした。アラスカ、カリフォルニア、アラバマ、ケベックのコヨーテには、オオカミの祖先がほとんど見られません。ミズーリ州、イリノイ州、フロリダ州のコヨーテは、5 ~ 10% のオオカミの祖先を示します。アカオオカミではオオカミの 40% とコヨーテの 60% の祖先があり、ヒガシシンリンオオカミではオオカミの 60% とコヨーテの 40% が、五大湖オオカミではオオカミの 75% とコヨーテの 25% でした。メキシコのオオカミと大西洋岸のオオカミではコヨーテの祖先が10%、太平洋岸とイエローストーンのオオカミでは5%、カナダ諸島のオオカミでは3%未満でした。3番目のイヌ科動物が北米のオオカミに似たイヌ科動物の混合に関与していたとしたら、その遺伝的特徴はコヨーテとオオカミで発見されたであろうが、発見されていない。
2018年、全ゲノム配列決定がイヌ属のメンバーの比較に使用されました。この研究は、コヨーテとハイイロオオカミの共通の祖先が、絶滅した未確認のイヌ科動物の幽霊集団と遺伝的に混ざったことを示しています。「ゴースト」イヌ科動物は遺伝的にドールに近く、アフリカの野生の犬が他のイヌ科動物種から分岐した後に進化しました。オオカミと比較したコヨーテの基礎的な位置は、コヨーテが未知の絶滅したイヌ科動物のミトコンドリアゲノムをより多く保持しているためであると提案されています。
亜種
2005 年の時点で、19 の亜種が認識されています。 コヨーテの地理的変異はそれほど大きくないが、全体として見ると、東部の亜種 ( C. l. thamnosおよびC. l. frustor ) は大型で濃い色の動物で、徐々に色が薄くなる。そして西と北に向かってサイズが縮小し(C. l. texensis、C. l. latrans、C. l. lestes、およびC. l. incolatus)、太平洋に向かって「黄土色」の色調(深いオレンジまたは茶色)が明るくなります。海岸(C. l. ochropus、C. l. umpquensis )、アリドアメリカ(C. l. microdon、C. l. mearnsi )でのサイズの縮小、メキシコと中央アメリカでは暗赤色と短い銃口への一般的な傾向人口。
亜種
三項権威 説明と画像
範囲
プレーンズ コヨーテC. l. ラトランスは亜種を指名する

セイ、1823 [同義語: C. l. ネブラセンシス(メリアム、1898)C.淡蒼球(メリアム、1897)]

最大の亜種。かなり淡い毛皮を持ち、大きな臼歯とカーナシアルを生やしています。
アルバータ州、マニトバ州、サスカチュワン州から南にニューメキシコ州とテキサス州パンハンドルまでの大平原
メキシコのコヨーテC. l. カゴッティ

CEH スミス、1839 年
C.lに似ています。半島ですが、より大きくて色が赤くなっています。耳は短く、歯は大きく、銃口は幅広です。
メキシコのオアハカ州、サンルイスポトシ州、プエブラ州、ベラクルス州
サン ペドロ マルティア コヨーテC. l. クレプティクス

エリオット、1903年
小さな亜種で、赤みがかった夏毛と短くて広い頭蓋骨を持っています。
バハ・カリフォルニア北部とカリフォルニア南西部
エルサルバドル コヨーテC. l. ディッキー

ネルソン、1932 年
大きな亜種で、C. l に相当します。サイズはレステスよりも小さいですが、歯は小さく、毛皮は暗めです。
当初はエルサルバドルのラ・ウニオンにあるゴアスコラン川の西3.2km(2マイル)にあるセロ・モゴテからのみ知られていました。 2013年1月には、その範囲を南方のパナマ南部まで拡大した。
サウスイースタンコヨーテC.l. イライラする人

ウッドハウス、1851
この亜種はC.l.に似ています。半島状ですが、より大きくて色が白く、耳は短く、銃口は長くなります。
カンザス州南東部および極東部、オクラホマ州、テキサス州、ミズーリ州、アーカンソー州
ベリーズ コヨーテC. l. ゴールドマニ

メリアム、1904 年
メキシコのコヨーテの中で最大のもので、C. l に近づきます。サイズはラトランスですが、銃口は短くなります。
グアテマラ国境に近いメキシコのチアパス州サンビセンテでのみ知られていますが、グアテマラ西部のコヨーテである可能性が
ホンジュラス コヨーテC. l. ホンジュレンシス

ゴールドマン、1936
小さな紅色の亜種で、粗くて薄い毛皮と広い頭蓋骨を持っています。
テグシガルパの北、アルカガの北東にある開けた土地でのみ知られている
デュランゴ コヨーテC. l. インパヴィドゥス

アレン、1903 年
このイヌ科動物はC.l に似ています。色はカゴッティですが、はるかに大きいです。
ソノラ州南部、チワワ最南西部、デュランゴ州西部、サカテカス州西部、シナロア州
ノーザンコヨーテC.l. インコラトゥス

ホール、1934 年
中型の亜種で、シナモン色の毛皮とC. l よりも凹んだ頭蓋骨を持っています。ラトランス。
アラスカ、ユーコン準州、ノースウェスト準州、ブリティッシュコロンビア州北部、アルバータ州北部の北方林
ティブロン島のコヨーテC. l. ジャメシ

タウンゼント、1912 年
C.lよりもはるかに薄いです。mearnsi、それはより重い歯、大きな頭蓋骨、そして長い耳を持っています。
ティブロン島
マウンテンコヨーテC.l. レステス

メリアム、1897
サイズと色はC.lに似ています。latrans、この亜種には大きな尾と耳が
ブリティッシュ・コロンビア州南部およびアルバータ州南東部、カスケード山脈以東のワシントン州、オレゴン州、カリフォルニア北部、モンタナ州西部、ワイオミング州、コロラド州(南東部を除く)、ネバダ州中北部、ユタ州中北部
ミアーンズのコヨーテC. l. マーンシ

メリアム、1897年[同義語: C. l. エスター(メリアム、1897)]

中くらいの大きさの耳、小さな頭蓋骨、小さな歯を持つ小さな亜種。その毛皮は豊かで明るい色です。フルーヴの色合いは非常に明るく、後足と前足を覆います。
コロラド州南西部、ユタ州とネバダ州の最南端、カリフォルニア州南東部、バハカリフォルニア州北東部、アリゾナ州、ニューメキシコ州のリオグランデ川以西、ソノラ州北部とチワワ州
ロウワー リオ グランデ コヨーテC. l. ミクロドン

メリアム、1897
小さな亜種で、小さな歯とかなり黒い毛皮を持っています。後足の上面は白っぽく、腹部には先端が黒い毛が散在しています。
テキサス州南部とタマウリパス州北部
カリフォルニア バレー コヨーテC. l. オクロプス

フォン・エシュショルツ、1829
C.lに似ています。ラトランスとC.l. lestesですが、小さく、暗く、より明るい色です。耳は大きく、頭蓋骨と歯は小さいです。
シエラネバダ以西のカリフォルニア
ペニンシュラ コヨーテC. l. 半島

メリアム、1897
C.lに似ています。サイズと特徴はオクロプスですが、毛皮はより暗く、より赤くなっています。尾の下側はC.l よりも黒いです。オクロプス、腹部には先端が黒い毛が多く
バハカリフォルニア
イースタンコヨーテC.l. 変数
ローレンスとボサート、1969 [代名詞: C. l. オリエンス、C.l. バージニエンシス]

これは、 C. lupus / C. lycaonとC. latransのハイブリッドです。ヒガシオオカミよりも小さく、狭い領土を保持しますが、典型的なニシオオカミよりも大きく、より広い行動範囲を保持します。
ニューイングランド、ニューヨーク、ニュージャージー、ペンシルベニア、オハイオ、ウェストバージニア、メリーランド、デラウェア、バージニア、およびカナダ東部のオンタリオ、ケベック、ニューブランズウィック、ノバスコシア、プリンスエドワード島、ニューファンドランド・ラブラドール州
テキサス プレーンズ コヨーテC. l. テセンシス

ベイリー、1905
C.lより小さい。latrans では、C. lに見られる豊かさに近い、より明るく、より豊かな毛皮を持っています。オクロプス、ただしC. l. テキセンシスには亜種の大きな耳がありません。
テキサス州の大部分、ニューメキシコ州東部、メキシコ北東部
ノースイースタンコヨーテC.l. タムノス

ジャクソン、1949 年
C.lとほぼ同じサイズ。latrans、またはそれより大きいが、色が暗く、より広い頭蓋骨を持っています。
サスカチュワン州中北部、マニトバ州(南西端を除く)、ケベック州東部から南部、ノースダコタ州南部から東部、ミネソタ州、アイオワ州、ミズーリ州(ミズーリ川の北)、ミシガン州、ウィスコンシン州、イリノイ州(最南部を除く) 、およびインディアナ州北部
ノースウェスト コースト コヨーテC. l. アンプクエンシス

ジャクソン、1949 年
小型の亜種で、黒っぽい赤みがかった毛皮、比較的小さな頭蓋骨、そして弱い歯列を持っています。
ブリティッシュコロンビア州、ワシントン州、オレゴン州の海岸
コリマ コヨーテC.l. 徹夜

メリアム、1897
C.lに似ています。半島ですが、より暗く、より広範囲に色が付いています。前腕には黒が多く、尾の下側には黒がありません(先端を除く)。
ハリスコ州南部からゲレーロ州までのメキシコの太平洋岸
ハイブリダイゼーション

メラニスティックコヨーテの色は、飼い犬で最初に生じた突然変異によるものです。
コヨーテは時々飼い犬と交尾し、口語的に「コイドッグ」として知られる交雑種を生み出すこともイヌとコヨーテの交尾周期は一致せず、コヨーテは通常イヌに対して敵対的であるため、野生ではこのような交配はまれである。交雑は通常、コヨーテが同種の動物が少なく、イヌが唯一の選択肢である地域に拡大している場合にのみ発生します。それでも、犬はコヨーテとつがいの絆を形成しないため、子の生存率は通常よりも低く、したがって子の飼育がより困難になります。飼育下では、F 1雑種(第一世代)は犬よりもいたずら好きで、子犬の頃は扱いにくい傾向があり、オオカミと犬の雑種よりも成熟したときの信頼度が低い。
雑種の外観はさまざまですが、通常はコヨーテの通常の特徴を保持しています。F 1ハイブリッドはイヌとコヨーテの中間的な形態となる傾向がありますが、F 2ハイブリッド (第 2 世代) はより多様です。F 1と F 2 のハイブリッドはどちらも、内気さと性的攻撃性の点で親のコヨーテに似ています。 雑種は繁殖力があり、4世代にわたって繁殖させることができます。 メラニスティックコヨーテの毛皮が黒いのは、飼い犬で最初に生じた突然変異によるものである。ニューファンドランド島の非アルビノホワイトコヨーテの個体群の色は、ゴールデンレトリバーから受け継いだメラノコルチン1受容体の突然変異によるものである。

雄のハイイロオオカミと雌のコヨーテの間で飼育下で生まれた
コイウルフの雑種
コヨーテは、特に北アメリカ東部で、さまざまな程度でオオカミと交雑しました。北アメリカ北東部のいわゆる「東部コヨーテ」は、おそらく北東部におけるハイイロオオカミと東部オオカミの絶滅の余波で発生し、その結果、コヨーテがかつてのオオカミの生息地に定着し、生き残ったオオカミの個体群と混ざることを可能にしました。この雑種はハイイロオオカミやヒガシオオカミよりも小さく、領土も狭いですが、典型的なニシコヨーテよりも体が大きく、より広い行動範囲を持っています。2010 年の時点では、ヒガシコヨーテの遺伝子構成はかなり均一であり、ヒガシオオカミやニシコヨーテからの影響は最小限です。
イースタンコヨーテの成体はウエスタンコヨーテよりも大きく、メスのイースタンコヨーテの体重はオスのウエスタンコヨーテよりも21%重いです。 生後35日までに身体的な違いがより明らかになり、東部のコヨーテの子は西部のコヨーテよりも脚が長い。歯の発育にも違いがあり、東部コヨーテでは歯の萌出が遅く、順序も異なります。大きさを除けば、ヒガシコヨーテはセイスタンコヨーテと物理的に似ています。4 つの色相の範囲は、ダークブラウンからブロンド、または赤みがかったブロンドまでありますが、最も一般的な相は灰色がかった茶色で、脚、耳、脇腹が赤みがかっています。
東部コヨーテと西部コヨーテの間には、攻撃性や喧嘩において大きな違いはありませんが、東部コヨーテは喧嘩が少なく、遊び好きである傾向が遊び行動に先立ってケンカが起こるウエスタンコヨーテの子犬とは異なり、イースタンコヨーテの子犬の間でのケンカは遊びが始まった後に起こります。東洋のコヨーテは、西洋のコヨーテよりもはるかに遅く、2歳で性的成熟に達する傾向が
ヒガシオオカミとレッドオオカミも、さまざまな程度のオオカミとコヨーテの交配の産物です。ヒガシオオカミはおそらく、親ハイイロオオカミ集団との大規模な戻し交雑とオオカミとコヨーテの混合の結果であったと考えられます。アカオオカミは、森林地帯の南東部でオオカミの個体数が減少していた時期に、オオカミとコヨーテの交配を余儀なくされ、また地元の親コヨーテ個体群と戻し交配が行われ、現生のアカオオカミのゲノムの約 75 ~ 80% が複製されて誕生した可能性がコヨーテ由来。
行動
社会的および生殖行動

ミアーンズのコヨーテ ( C. l. mearnsi ) の子犬が遊んでいます。

イエローストーン国立公園のコヨーテの群れ
コヨーテは、ユーラシアのキンイロジャッカルと同様に群生しますが、オオカミのようなより社会的なイヌ科の種ほど同種に依存しません。これは、コヨーテが後者の種のように大型の獲物を専門に狩らないためであると考えられます。コヨーテの群れの基本的な社会単位は、生殖能力のあるメスを含む家族です。しかし、血縁関係のないコヨーテが仲間を求めたり、単独で攻撃するには大きすぎる獲物を仕留めるために力を合わせることもこのような「家族以外の」群れは一時的なものにすぎず、独身男性、生殖能力のない女性、および亜成人の若者で構成されている場合がメスが発情期を迎える真冬に家族が形成されます。つがいの結合は、実際の交尾が行われる 2 ~ 3 か月前に起こることが
交尾の結びつきは5 ~ 45 分間続くことが発情期に入ったメスは、匂いのマーキングと頻度の増加によって雄鳴きをすることでオスを誘引する。発情期の 1 匹のメスは最大 7 匹の生殖能力のあるオスを引き寄せることができ、そのオスは 1 か月間も追いかけることができます。オス同士の争いが起こることもありますが、メスが交尾相手を選んで交尾すると、拒絶されたオスは介入せず、他の発情したメスを見つけると次の行動に移ります。一夫一婦制と重婚制の両方の交配を行うことが知られているオオカミとは異なり、コヨーテは、コヨーテ密度が高く、餌が豊富な地域であっても、厳密に一夫一婦制である。
交尾に失敗したメスは、姉妹や母親の子育てを手伝ったり、次に交尾できるようになるまで兄弟に加わったりすることがその後、新しく交尾したペアは縄張りを確立し、自分の巣穴を建設するか、放棄されたアナグマ、マーモット、またはスカンクアースを掃除します。妊娠中、オスは頻繁に単独で狩りをし、メスのために食べ物を持ち帰ります。メスは乾いた草やお腹から抜いた毛皮で巣穴を覆うことも妊娠期間は63日で、平均産児数は6匹だが、その数はコヨーテの個体数密度と餌の豊富さによって変動する。
コヨーテの子は巣穴、木の洞、または棚の下で生まれ、出生時の体重は 200 ~ 500 g (0.44 ~ 1.10 ポンド) です。彼らは禁酒性であり、生後10日間は完全にミルクに依存します。切歯は約12日で生え、犬歯は16日で、第二小臼歯は21日で生えてきます。10日後に目が開き、その時点までに子犬はますます動けるようになり、20日で歩き、6週齢で走り始めます。 。親は、12 ~ 15 日後に吐き出した固形食品を子犬の食事に補給し始めます。乳歯が完全に機能する生後4~6週目までに、子犬にはマウス、ウサギ、有蹄類の死骸などの小さな餌が与えられ、2か月後には授乳量が着実に減少します。
オオカミの子とは異なり、コヨーテの子は遊び行動に入る前に(喧嘩ごっこではなく)真剣に戦い始めます。一般的な遊び行動には、コヨーテの「ヒップスラム」が含まれます。生後3週間までに、コヨーテの子はオオカミの子よりも抑制力なく互いに噛みつきます。生後4~5週目までに、子犬は優位な階層を確立し、その頃には喧嘩するよりも遊ぶ傾向が強くなります。オスは子犬に餌を与え、毛繕いをし、見守るという積極的な役割を果たすが、子犬が完全に乳離れする前にメスが行方不明になると、子犬を放棄する。巣穴は6月から7月までに放棄され、子犬は親の後を追って縄張りを巡回したり狩りをしたりする。子犬は8月に家族のもとを離れることがありますが、それよりずっと長く残ることも子犬は8か月で成犬の寸法に達し、1か月後には成犬の体重が増加します。
縄張りと避難行動
個々の採餌領域のサイズは 0.4 ~ 62 km 2 (0.15 ~ 24 平方マイル) であり、特定の地域におけるコヨーテの一般的な集中度は、餌の豊富さ、適切な巣穴の場所、同種および他の捕食者との競争によって異なります。コヨーテは一般にデニングシーズン以外は自分の縄張りを守らず、オオカミほど侵入者に対して攻撃的ではなく、通常は侵入者を追いかけたりスパーリングをしたりするが、殺すことはめったにない。食糧不足の時にはコヨーテ間の争いが起こることがある。コヨーテは足を上げて放尿したり地面を引っ掻いたりして自分の縄張りをマークする。
オオカミと同様、コヨーテは妊娠や子育ての際に巣穴(通常は他の種の人のいない穴)を利用しますが、屋外のヨモギの下で出産することもコヨーテの巣穴は、渓谷、流域、クーリー、堤防、岩の断崖、または平地にいくつかの巣穴は、放棄された自家農園の小屋、穀物箱、排水管、線路、空洞の丸太、藪、アザミの下で発見されています。巣穴は子が生まれるまでメスによって継続的に掘られ、掃除されます。巣穴が荒らされたり、ノミが蔓延したりした場合、子犬は別の巣穴に移動されます。コヨーテの巣穴には、主室から分岐したいくつかの入り口や通路がある場合が 1 つの巣穴を何年も使用できます。
狩猟と採食行動
嗅覚は狩猟にとって非常に重要であるというのが一般的なコンセンサスですが 、嗅覚、聴覚、視覚の合図の役割を実験的に調査した 2 つの研究では、アカギツネとコヨーテでは視覚の合図が狩猟にとって最も重要であることがわかりました。。

獲物に飛びつくコヨーテ。
大きな獲物を狩るとき、コヨーテはペアまたは小さなグループで行動することがよく大型の有蹄動物を仕留められるかどうかは、雪の深さや地殻の密度などの要因によって決まります。通常、若い動物はそのような狩猟に参加することを避け、繁殖ペアが作業のほとんどを行うのが一般的です。コヨーテは大きな獲物を追いかけ、典型的には動物のハムストリングを起こし、その後獲物が落ちるまで嫌がらせをする。他のイヌ科動物と同様に、コヨーテは余分な食べ物を隠します。コヨーテはネズミほどの大きさのげっ歯類を飛びついて捕まえるが、ジリスは追いかける。コヨーテは大きな群れで生きることもありますが、小さな獲物は通常単独で捕らえられます。
コヨーテはペアでヤマアラシを殺し、前足を使ってげっ歯類を仰向けにし、柔らかい下腹部を攻撃することが観察されている。ヤマアラシをうまく捕食できるのは年老いた経験豊富なコヨーテだけであり、若いコヨーテによる何度も捕食の試みが行われ、獲物の羽根で怪我をすることになります。コヨーテは、おそらく食べ物に対する所有権を主張するために、食べ物の上に放尿することが 最近の証拠は、おそらく人間を避けるために、少なくとも一部のコヨーテが狩猟においてより夜行性になったことを示している。
コヨーテは時折、アメリカアナグマと相利的な狩猟関係を築き、げっ歯類の獲物を掘り出す際に互いに助け合うことがコヨーテの中には、アナグマの仲間に頭を横たわったり、抵抗せずに顔をなめたりする姿が観察されているため、2種の関係は時として見かけ上の「友情」に近いかもしれない。コヨーテとアナグマの友好的な交流は、メキシコで発見された西暦 1250 ~ 1300年の壺に両者の関係が描かれていることから、コロンブス以前の文明には知られていました。
食べ残し、ペットフード、動物の糞便は、コヨーテをゴミ箱に引き寄せる可能性が
コミュニケーション

コヨーテの遠吠え
夜に吠えるコヨーテの群れ
ボディランゲージ
社交的であると同時に単独で行動する動物でもあるコヨーテの視覚と音声のレパートリーの多様性は、単独で行動するキツネと高度に社交的なオオカミの中間となります。コヨーテの攻撃的な行動は、オオカミや犬よりもキツネの行動に似ている 。攻撃的なコヨーテは背中を反らせ、尻尾を下げます。「お辞儀」に続いて「跳躍遊び」を行うことで遊びの行動を促すイヌとは異なり、コヨーテの遊びは、お辞儀に続いて頭を左右に屈曲させ、一連の「回転」を行うことで構成される。 」と「ダイブ」。コヨーテも犬と同じように遊び相手の首筋を噛むことがありますが、通常は低い位置に近づき、上向きに噛みつきます。
子犬は性別に関係なく互いに戦いますが、成犬の場合、攻撃性は通常、同性のメンバーに限定されます。戦闘員は尻尾を振り、顎を開いてうなり声を上げながら互いに近づきますが、戦いは通常静かに行われます。オスは垂直姿勢で戦う傾向がありますが、メスは4本の足すべてで戦います。メス同士の喧嘩は、メスが相手の前足、喉、肩を掴むため、オス同士の喧嘩よりも深刻になる傾向が
発声
叫ぶコヨーテ
コヨーテは「北米のすべての野生哺乳類の中で最も声が大きい」と言われています。 その声の大きさと鳴き声の範囲が、「吠える犬」を意味する二項名Canis latransの由来となった。大人のコヨーテでは少なくとも 11 種類の異なる鳴き声が知られています。これらの音は、苦痛音と警報音、挨拶音、接触音の 3 つのカテゴリに分類されます。最初のカテゴリの発声には、ウーウー、うなり声、ハフ声、吠え声、吠える声、叫び声、および高周波の鳴き声が含まれます。ワンワン音は低強度の威嚇や警報として使用され、通常は巣穴の近くで聞こえ、子犬がすぐに巣穴に逃げるよう促します。
うなり声は近距離での威嚇として使用されますが、雄同士で遊んだり交尾したりする子犬の間でも聞こえます。ハフとは、空気を急速に吐き出すことによって生成される、強度の高い脅威の発声です。吠え声は、長距離の威嚇の声と警報の鳴き声の両方に分類できます。樹皮の遠吠えも同様の機能を果たす可能性が叫び声は服従の兆候として発せられますが、高周波の鳴き声は、従属者の服従を認識する支配的な動物によって生成されます。挨拶の発声には、低周波の鳴き声、「わーわーわー」、集団でのイーップの遠吠えなどが低周波の鳴き声は従順な動物によって発せられ、通常は尻尾を振ったり、銃口をかじったりします。
「wow-oo-wow」として知られるサウンドは、「挨拶の歌」として説明されています。集団イップの遠吠えは、2 人以上の群れのメンバーが再会するときに発せられ、複雑な挨拶儀式の最後の行為である可能性がコンタクトコールには、単独の遠吠えとグループの遠吠え、および前述のグループイップの遠吠えが含まれます。孤独な遠吠えはコヨーテの最も象徴的な鳴き声であり、群れから離れた一匹の個体の存在を知らせる目的があると考えられます。グループの遠吠えは、グループ yip の遠吠えの代替として、また単独の遠吠え、グループ yip の遠吠え、またはグループ yip の遠吠えに対する応答として使用されます。
エコロジー編集
生息地

サンフランシスコ、バーナルハイツの都会的なコヨーテ
オオカミやクーガーがほぼ絶滅する前は、コヨーテはバイソン、プロングホーン、ヘラジカ、その他の鹿が生息する草原に最も多く生息しており、特にプレーリードッグの生息する草の短い地域でよく生息していましたが、世界でも同様にコヨーテが生息していました。ヨモギやジャックウサギが生息する半乾燥地帯、あるいはサボテン、カンガルーネズミ、ガラガラヘビが生息する砂漠など。オオカミと直接競争していない限り、コヨーテはソノラ砂漠から隣接する山々の高山地域、あるいはアルバータ州の平野や山岳地帯まで生息していました。オオカミの絶滅により、コヨーテの生息域はグアテマラの熱帯地方とアラスカの北斜面の荒廃した森林を含むように拡大しました。
コヨーテは 1 日に約 5 ~ 16 キロメートル (3 ~ 10 マイル) を歩き、多くの場合、伐採道路や小道などの小道に沿って歩きます。冬には氷に覆われた川を移動ルートとして使用する場合が多くの場合、薄明薄膜であり、日中よりも夕方から夜の初めにかけて活動的になります。しかし、都市部ではコヨーテは夜行性であることが知られており、人間との遭遇を避ける可能性が高い。多くのイヌ科動物と同様に、コヨーテは泳ぎが上手で、水面を少なくとも0.8キロメートル(0.5マイル)移動できると報告されている。
ダイエット

ワイオミング州シードスカディー国立野生動物保護区で、道路に落ちたプロングホーンの破片を持つコヨーテ
コヨーテは、生態学的にはユーラシアのキンイロジャッカルに相当する北米の動物です。同様に、コヨーテは食べ物の選択において非常に多様性がありますが、主に肉食であり、食事の90%が肉で構成されています。獲物の種には、バイソン(主に死肉として)、オジロジカ、ラバジカ、ヘラジカ、ヘラジカ、オオツノヒツジ、プロングホーン、ウサギ、ノウサギ、齧歯動物、鳥類(特にガリ目、ロードランナー、若い水鳥、ハトおよびハト)、両生類が含まれます。 (ヒキガエルを除く)、トカゲ、ヘビ、カメ、魚、甲殻類、昆虫。コヨーテは、トガリネズミ、モグラ、ドブネズミなどの動物がその数に比例して食事に含まれないため、狙う獲物にうるさい可能性が
陸生動物、および/またはジリスやその関連種(マーモット、プレーリードッグ、シマリス)、ハタネズミ、ポケットホリネズミ、カンガルーネズミ、その他の地上性のげっ歯類などの穴を掘る小型哺乳類は、特に単独のコヨーテにとって、非常に一般的な餌となります。 特定の主要な哺乳動物の獲物の例としては、ワタオウサギ、サーモンジリス、およびシロアシネズミが挙げられる。さらに珍しい獲物には、漁師、ツキノワグマの子、タテゴトアザラシ、ガラガラヘビなどがある。コヨーテは主に食用としてガラガラヘビを殺しますが、巣穴で子を守るためにも、ヘビが伸びるまでからかってから頭を噛んだり、ヘビを折ったり揺すったりして殺します。コヨーテに捕らえられる鳥の大きさは、スラッシャー、ヒバリ、スズメから野生の七面鳥の成鳥、そしてまれに、抱っこしている成鳥の白鳥やペリカンまで多岐にわたります。
コヨーテは群れやつがいで行動する場合、通常単独で捕食するよりも大きな獲物、たとえば重さ10kg(22ポンド)を超えるさまざまな獲物にアクセスできる可能性が 場合によっては、コヨーテの群れがオドコイレウスジカの成体、カウヘラジカ、プロングホーン、野生の羊などのはるかに大きな獲物を送り込んだこともあるが、これらの動物の若い子鹿、子牛、子羊はかなり頻繁に捕獲される。パック、家畜羊および家畜牛も同様です。場合によっては、コヨーテは体重 100 ~ 200 kg (220 ~ 440 ポンド) 以上の獲物を仕留めることも野生のシカなどの成体有蹄動物の場合、衰弱している、雪や氷の中に閉じ込められている、冬で衰弱している、妊娠しているなどの弱い立場にあるときに搾取されることがよくありますが、警戒心が弱い家畜の有蹄類はより簡単に搾取される可能性が
コヨーテは新鮮な肉を好みますが、機会があれば肉をあさります。食べる昆虫、果物、草を除くと、コヨーテは毎日推定 600 g (1.3 ポンド)、または年間 250 kg (550 ポンド) の餌を必要とします。コヨーテは同種の動物の死骸を容易に共食いし、コヨーテの脂肪はコヨーテハンターによっておびきや毒餌としてうまく利用されてきた。コヨーテの冬の食事は主に大きな有蹄動物の死骸で構成されており、植物はほとんど含まれげっ歯類の獲物は、春、夏、秋に重要性が増します。
コヨーテは、イチゴ、ブラックベリー、ブルーベリー、サルサパリラ、桃、梨、リンゴ、ウチワサボテン、シャポー、柿、落花生、スイカ、マスクメロン、ニンジンなど、さまざまな農産物を食べます。冬から早春にかけて、コヨーテは緑の小麦の葉などの草を大量に食べます。綿餅、大豆粕、家畜の糞、豆、トウモロコシ、小麦、ソルガムなどの栽培穀物などの珍しい物を食べることも
カリフォルニア沿岸では、コヨーテは現在、祖先よりも高い割合で海洋性食品を摂取していますが、これはこの地域からハイイログマが絶滅したためと考えられています。デスバレーでは、春の開花期にコヨーテがスズメガの 毛虫や甲虫を大量に食べることが
敵と競争相手

コヨーテとハイイロオオカミの比較イラスト

若いクーガーを追い詰めるマウンテンコヨーテ ( C. l. lestes )
コヨーテとハイイロオオカミの生息域が重なる地域では、オオカミによる干渉競争と捕食が、局所的なコヨーテ密度を制限しているという仮説が立てられている。コヨーテの生息域は、オオカミの絶滅に続いて 19 世紀から 20 世紀にかけて拡大しましたが、 1940 年代にオオカミがロイヤル島に定着した後、コヨーテはロイヤル島で絶滅に追い込まれました。両種が共存するイエローストーン国立公園で行われたある研究では、 1990年代のオオカミの再導入後、ラマー川渓谷のコヨーテの個体数は39%減少したが、グランドティトン国立公園のオオカミ生息地域のコヨーテの個体数は減少したと結論づけた。存在しない地域に比べて 33% 低い。 コヨーテはオオカミを尾行して獲物を食べることが知られているが、オオカミは近くにコヨーテを許容しないことが観察されている。
地域によってはコヨーテがクーガーと競合することもシエラネバダ東部では、コヨーテがミュールジカをめぐってクーガーと争っています。クーガーは通常、コヨーテと競争して優勢であり、時折コヨーテを殺す可能性があるため、キツネやボブキャットなどの小型肉食動物に対するコヨーテの捕食圧力が軽減されます。殺されたコヨーテは食べられないこともあり、おそらくこれらが競合的な種間相互作用を構成していることを示しているが、クーガーもコヨーテを食べるという確認された例が複数ある。 メキシコ北東部では、コヨーテに対するクーガーの捕食が急速に続いているが、明らかに生息地の利用方法が異なるため、同所性ジャガーの獲物の範囲にコヨーテは含まれていない。
ハイイロオオカミとクーガーを除けば、成体のコヨーテが捕食されることは比較的まれですが、他の複数の捕食者が時折脅威になる可能性が成体のコヨーテがアメリカクロクマとハイイログマの両方、アメリカワニ 、大型のカナダオオヤマネコ 、イヌワシによって捕食されたケースもある。殺害現場や死肉では、コヨーテは、特に単独で行動する場合、オオカミ、クーガー、クマ、クズリ、そして通常は常にではないがワシ(ハゲワシや金ワシ)に支配される傾向がある。このような、より大きく、より強力で、より攻撃的な捕食者が共有の餌場にやってくると、コヨーテは戦おうとするか、他の捕食者が終わるまで待つか、時には殺し合いをすることもありますが、次のような重大な危険があった場合には、オオカミや大人のクーガーがいると、コヨーテは逃げる傾向が
コヨーテが健康な大人のアカギツネを殺すことはめったになく、彼らと一緒に餌を食べたり巣穴を掘ったりする様子が観察されていますが、罠にかかったキツネを殺すことはよくコヨーテはキツネのキットを殺す可能性がありますが、これは主要な死亡原因ではありません。南カリフォルニアではコヨーテが頻繁にハイイロキツネを殺しており、これらの小型のイヌ科動物はコヨーテ密度の高い地域を避ける傾向がある。
一部の地域では、コヨーテはボブキャットと生息域を共有しています。コヨーテ密度が高い地域ではボブキャットの個体数が減少する傾向にありますが、これら 2 つの同様のサイズの種が物理的に互いに対峙することはほとんどありません。しかし、いくつかの研究ではコヨーテとボブキャットの間の干渉競合が実証されており、すべての場合においてコヨーテが相互作用を支配していた。 複数の研究者 はコヨーテがボブキャットを殺す例を報告したが、ボブキャットがコヨーテを殺すことはより稀である。コヨーテは、中型の獲物に使用されるのと同様の噛みつき振り方を使用してボブキャットを攻撃します。コヨーテは、単独の個体でも集団でも、時折ボブキャットを殺すことが知られています。ほとんどの場合、ボブキャットは成体雌や幼体などの比較的小さな標本でした。
未知の数のコヨーテによる、成体の雄のボブキャットに対するコヨーテ攻撃が発生しました。カリフォルニアでは、コヨーテとボブキャットの個体数は、異なる生息地の種類間で負の相関関係はありませんが、コヨーテによる捕食はボブキャットの重要な死亡原因となっています。生物学者のスタンリー・ポール・ヤングは、わな猟のキャリアを通じて、捕獲したボブキャットをコヨーテに殺されそうになったことが一度も成功したことがないと指摘し、コヨーテがボブキャットを木の上まで追いかけた2件の事件について書いた。コヨーテは時折カナダオオヤマネコを直接殺し、獲物、特にカンジキウサギをめぐって彼らと争うことが記録されている。アルバータ州中央部を含む一部の地域では、コヨーテが少ない地域ではオオヤマネコがより多く生息しているため、カンジキウサギの入手可能性よりもコヨーテとの相互作用の方がオオヤマネコの個体数に影響を与えているようだ。
範囲
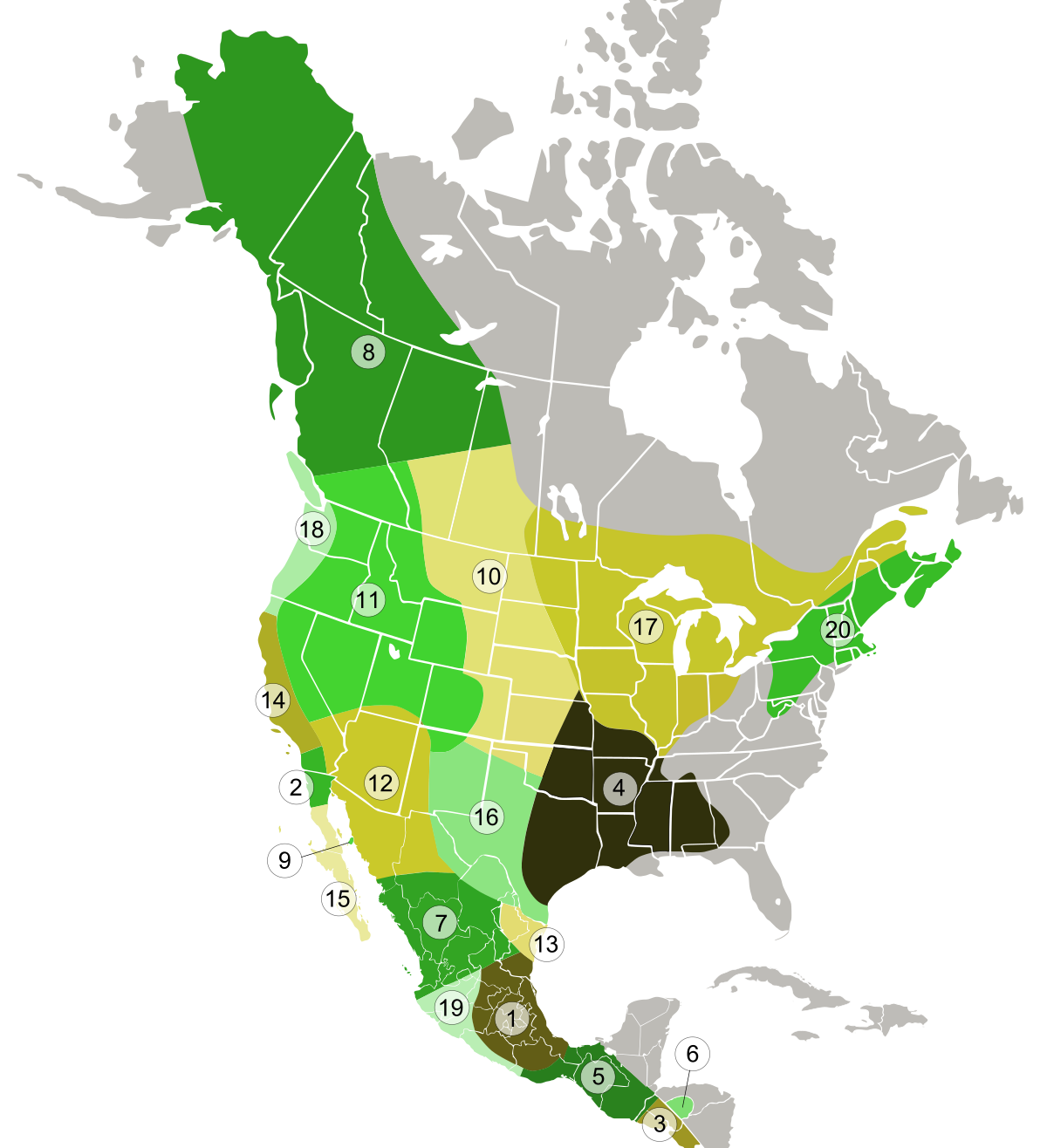
1978 年の時点でのコヨーテ亜種の範囲: (1) メキシコ コヨーテ、(2) サン ペドロ マルティア コヨーテ、(3) エルサルバドル コヨーテ、(4) サウスイースタン コヨーテ、(5) ベリーズ コヨーテ、(6) ホンジュラス コヨーテ、(7) ) デュランゴ コヨーテ、(8) ノーザン コヨーテ、(9)ティブロン島コヨーテ、(10)平原コヨーテ、(11) マウンテン コヨーテ、(12)ミアーンズ コヨーテ、(13) ローワー リオグランデ コヨーテ、(14) カリフォルニア バレー コヨーテ、(15)半島コヨーテ、(16)テキサス平原コヨーテ、(17)北東部コヨーテ、(18)北西海岸コヨーテ、(19)コリマコヨーテ、(20)東部コヨーテ

過去1万年にわたるコヨーテの拡大

1900年以来数十年にわたるコヨーテの拡大
コヨーテは北米全域に広範囲に生息し、生息数が多いため、国際自然保護連合(IUCN)によって軽度懸念事項に指定されています。コロンブス以前のコヨーテの生息域は、北米の南西部と平原地域、メキシコの北部と中部に限定されていた。19 世紀までにこの種は北と東に拡大し、土地転換とオオカミの絶滅と時を同じくして 1900 年以降さらに拡大しました。この時までに、その範囲は、隣接する米国とメキシコの全土を含む北米大陸全体を網羅し、南は中央アメリカ、北はカナダとアラスカの大部分に及んでいました。この拡大は進行中であり、現在この種は北緯 8 度 (パナマ) から北緯 70 度 (アラスカ北部) の間の地域の大部分を占めています。
かつては、コヨーテは森林伐採によって拡大を助けられ、メキシコ南部や中央アメリカに最近移住したものであると広く信じられていましたが、更新世と完新世初期の記録、さらには先コロンブス期と初期のヨーロッパ植民地化の記録は、この動物が森林伐採によって拡大したことを示しています。近代よりずっと前からこの地域に存在していました。熱帯雨林への牛の放牧地拡大に続き、1970 年代後半にコスタリカ南部で、1980 年代初頭にパナマ北部で生息域の拡大が起こりました。
コヨーテはベリーズ北部の生息地がコヨーテにとって好ましい環境であるため、近い将来ベリーズ北部に出現すると予測されている。パンアメリカン・ハイウェイによってダリエン溝が塞がれた場合、パナマ地峡を通って南アメリカに進出する可能性についての懸念が提起されている。この懸念は 2013 年 1 月に部分的に確認され、この種がパナマ運河を越えたパナマ東部のチェポ地区で記録されました。
2017年の遺伝子研究では、コヨーテはもともと米国東部の地域では発見されていなかったことが示唆されています。1890 年代以降、密林が農地に変わり、オオカミの駆除が大規模に実施されたため、コヨーテが分散するニッチが残されました。遺伝的に異なるコヨーテの 2 つの集団から 2 つの大きな分散がありました。北東部への最初の大規模な分散は、20 世紀初頭に、グレート プレーンズ北部に生息するコヨーテから起こりました。これらは五大湖地域北部とカナダ南部を経由してニューイングランドに、そして五大湖地域南部を経由してペンシルベニアに伝わり、1940年代にニューヨークとペンシルベニアで合流した。
これらのコヨーテは、残存するハイイロオオカミやヒガシオオカミの個体群と交配し、コヨーテの遺伝的多様性を高め、新しいニッチへの適応を助けた可能性が南東部への 2 番目の大規模な拡散は 20 世紀半ばにテキサス州から到来し、1980 年代にカロライナ州に到達しました。これらのコヨーテは、アカオオカミが野生で絶滅した1970年代より前に、残存していたアカオオカミの個体群と交雑しており、それがコヨーテの遺伝的多様性をさらに高め、この新たなニッチへの適応にも役立っている可能性がある。これら 2 つの主要なコヨーテの分散は両方とも急速な個体数増加を経験しており、中部大西洋岸に沿って会合すると予測されています。この研究は、コヨーテの長距離分散、局所個体群からの遺伝子流出、急速な個体数増加が相互に関連している可能性があると結論付けている。
病気と寄生虫

肉腫疥癬に苦しむカリフォルニア バレー コヨーテ ( C. l. ochropus )
北米の大型肉食動物の中で、コヨーテはおそらく最も多くの病気や寄生虫を媒介しますが、これはおそらくその広範囲で多様な食性によるものと考えられます。 コヨーテに感染することが知られているウイルス性疾患には、狂犬病、犬ジステンパー、犬伝染性肝炎、4種類の馬脳炎、口腔乳頭腫症などが1970 年代後半までに、コヨーテでの深刻な狂犬病の発生は 60 年以上問題ではなくなりましたが、1 ~ 5 年おきに散発的に発生しました。ジステンパーは野生では多くの子犬を死に至らしめますが、一部の個体は感染しても生き残ることができます。細菌性疾患である野兎病は、ダニの刺咬やげっ歯類やウサギ目などの獲物を介してコヨーテに感染し、子犬にとっては致命的となる可能性が
コヨーテは毛包虫と疥癬の両方に感染する可能性があり、後者が最も一般的です。コヨーテではダニの侵入はまれで偶発的ですが、ダニの侵入はより一般的で、季節のピークは地域によって異なります(北西部では5月から8月、アーカンソー州では3月から11月)。コヨーテにシラミが寄生することはほとんどありませんが、ノミはコヨーテの幼少期から寄生しますが、重篤な病気というよりも炎症の原因となる可能性がPulex simulans はコヨーテに寄生する最も一般的な種ですが、Ctenocepharides canis はコヨーテと犬 (主な宿主) が同じ地域に生息する場所でのみ発生する傾向がコヨーテが吸虫の宿主となることはめったにありませんが、それでもコヨーテ、特に死亡率 90% の病気であるサケ中毒病に感染する可能性のあるナノフィエトゥス サルミンコーラに深刻な影響を与える可能性が結腸吸虫もコヨーテに感染する可能性が
サナダムシは、調査したすべてのコヨーテの 60 ~ 95% に寄生していることが記録されています。コヨーテに寄生する最も一般的な種は、ワタオウサギを中間宿主として使用する有鉤条虫と有鉤条虫です。コヨーテの中で知られている最大の種はT. hydatigenaで、感染した有蹄動物からコヨーテに侵入し、体長 80 ~ 400 cm (31 ~ 157 インチ) まで成長することがかつては主にオオカミに限定されていましたが、エキノコッカス・グラヌロススは、コヨーテがかつてのオオカミの生息地に定着し始めて以来、コヨーテにまで拡大しました。
コヨーテに最も頻繁に発生するアスカロイド回虫はトキサスカリス・レオニナで、コヨーテの小腸に常在し、宿主の食欲を高める以外に悪影響はありません。アンシロストマ属の鉤虫は、コヨーテの生息範囲全体に寄生し、特に湿気の多い地域で蔓延します。テキサス州沿岸などの湿気の多い地域では、コヨーテはそれぞれ最大 250 匹の鉤虫を運ぶことが吸血性のA.カニナムは、失血や肺うっ血によってコヨーテにダメージを与えるため、特に危険です。生後 10 日の子犬が、わずか 25 匹のA. caninum虫の宿主になると死亡する可能性が
人間との関係
詳細情報:アーバンコヨーテ
民間伝承や神話では
詳細は「コヨーテ (神話)」を参照

エドワード・S・カーティスのインド時代の昔、カヌーで漕ぐコヨーテ
コヨーテは、一部のネイティブアメリカン、特に南西部と平原地域のいくつかの国の民話の中でトリックスターの人物およびスキンウォーカーとして登場し、実際のコヨーテまたは人間の姿を交互にとります。他のトリックスターの人物と同様に、コヨーテは、欺瞞とユーモアを通じて社会的慣習に反逆するピカレスクのヒーローとして行動します。ハリスなどの民俗学者は、コヨーテがその知能と順応性の高さからトリックスターとみなされるようになったと考えている。ヨーロッパによるアメリカ大陸の植民地化後、英米ではコヨーテは臆病で信頼できない動物として描かれている 。公共イメージが根本的に改善されたハイイロオオカミとは異なり、コヨーテに対する英米の文化的態度は依然として否定的なものが多い。
マイドゥの創造物語の中で、コヨーテは仕事、苦しみ、死を世界に紹介します。ズニ族の言い伝えでは、コヨーテはカチナから光を盗んで世界に冬をもたらすとされています。チヌーク、マイドゥ、ポーニー、トホノ・オーダム、ユートはコヨーテを創造主の仲間として描いています。トホノ・オーダムの洪水物語では、コヨーテが人類を滅ぼす世界的な大洪水からモンテスマを生き延びるのを助けます。創造主が人類を創造した後、コヨーテとモンテズマは人々に生き方を教えます。クロウの創造物語では、コヨーテ老人が創造主として描かれています。ディネ創造物語では、コヨーテは最初の男と最初の女とともに第一世界に存在していましたが、別のバージョンでは第四世界で作成されました。ナバホ族のコヨーテは世界に死をもたらし、死がなければ人間が多すぎてトウモロコシを植える余地がなくなると説明しています。

コヨーテの戦士を描いた
テオティワカンのアテテルコの壁画
スペインがアステカ帝国を征服する前、コヨーテはメソアメリカの宇宙論で重要な役割を果たしました。コヨーテは古典時代の テオティワカンにおいて軍事力を象徴しており、戦士たちはその略奪的な力を呼び起こすためにコヨーテの衣装を着ていました。この種は、古典的アステカの統治が終わるまでの何世紀にもわたって、中央メキシコの戦士崇拝と結びつき続けました。
アステカ神話では、ダンス、音楽、肉欲の神であるウエウェコヨトル(「古いコヨーテ」の意味)は、いくつかの写本でコヨーテの頭を持つ男性として描かれています。彼は愛の女神ソチケツァルを誘惑して世界に戦争をもたらした女たらしとして描かれることもある。碑文学者デイヴィッド・H・ケリーは、ケツァルコアトル神はアステカ以前のウト・アステカ神話におけるコヨーテの描写にその起源があると主張し、コヨーテは創造者、誘惑者、トリックスター、文化英雄として人類の「兄」として描かれている。明けの明星へ。
人間への攻撃
詳細は「コヨーテによる人間への攻撃」を参照

コヨーテに餌を与えることを思いとどまらせる標識。コヨーテが人間の存在に慣れてしまい、攻撃の可能性が高まる可能性がある
コヨーテが人間を襲うことはまれで、コヨーテは比較的小さいため重傷を負うことはほとんどありませんが、特にカリフォルニアではその頻度が高まっています。確認された致命的な攻撃は2件のみで、1件はカリフォルニア州グレンデールの3歳のケリー・キーン君に対するもので 、もう1件はカナダのノバスコシア州のテイラー・ミッチェルという19歳の少年に対するものである。 2006年3月までの30年間に、米国では少なくとも160件の攻撃が発生し、そのほとんどがロサンゼルス郡地域で発生した。米国農務省(USDA)野生生物局、カリフォルニア州魚類猟獣局、その他の情報源からのデータによると、1988 年から 1997 年の期間に 41 件の攻撃が発生しましたが、1998 年から 2003 年までに 48 件の攻撃が確認されました。これらの事件の大部分は、郊外と原野の境界付近の南カリフォルニアで発生しました。
田舎の人々によるコヨーテへの嫌がらせがなくなったため、都市部のコヨーテは人間に対する恐怖心を失いつつあり、人々が意図的または非意図的にコヨーテに餌を与えることでさらに状況が悪化しています。このような状況では、一部のコヨーテは人間に対して攻撃的に行動し始め、ジョギングや自転車に乗る人を追いかけたり、犬の散歩をしている人に対峙したり、小さな子供にストーキングしたりするようになりました。これらの地域の非狂犬性コヨーテは、ほとんどが10歳未満の小さな子供を狙うこともありますが、大人が噛まれた例も
このような攻撃に関するメディア報道は一般に、問題の動物を単なる「コヨーテ」と特定しているが、東部コヨーテの遺伝学の研究によると、ペンシルベニア、ニューヨーク、ニューイングランド、カナダ東部を含む北アメリカ北東部での攻撃に関与した動物は、コヨーテの可能性があることが示されている。実際には、完全なコヨーテではなく、 Canis latransとC. lupusのハイブリッドであるコイウルフでした。
家畜やペットの捕食

犬と対峙するコヨーテ
2007 年の時点で、コヨーテは北アメリカ西部で最も多く存在する家畜捕食者であり、羊、ヤギ、牛の大部分を失っています。例えば、国立農業統計局によると、2004 年に捕食による羊の死亡数 224,000 頭のうち 60.5% がコヨーテによるものでした。 2004 年に死亡した羊の総数は、米国の羊と子羊の総頭数の 2.22% を占め、国立農業統計局 USDA の報告書によれば、2005 年 7 月 1 日の時点でそれぞれ合計 466 万頭と 780 万頭であった。
コヨーテの個体数は通常、オオカミの個体数の何倍も多く、より広範囲に分布しているため、コヨーテは全体としてより多くの捕食損失を引き起こします。米国政府職員は、家畜を守るために毎年約9万頭のコヨーテを定期的に射殺、毒殺、罠にかけて殺害している。 2005年に行われたアイダホ州の国勢調査では、コヨーテ個体はオオカミ個体よりも家畜を攻撃する可能性が5%高いことが示された。ユタ州では、2017年6月30日に終了する会計年度に、総額50万ドルを超える報奨金のために11,000頭以上のコヨーテが殺された。
家畜の番犬は捕食者を積極的に撃退するために一般的に使用されており、柵で囲まれた牧草地と放牧地の両方でうまく機能しています。米国の羊生産者を対象とした1986年の調査では、82%が犬の使用が経済的資産であると報告した。
牛の自然な保護傾向を高めることを含む牛の再野生化は、コロラド州立大学のテンプル・グランディンによって議論されたコヨーテの防除方法です。この方法は、牛の群れを牧場で出産させ、一年中開いた牧草地で牛を放牧する生産者の間で人気を集めている。

典型的な喉を押さえて家畜の羊を捕まえるコヨーテ
コヨーテは通常、大人のヒツジやヤギを攻撃するときに顎のすぐ後ろと耳の下の喉を噛み、窒息によって死亡することがよく通常、失血は二次的な死因です。子牛や羊の毛をたっぷりと被った羊は、脇腹や後肢を攻撃してショックや失血を引き起こして死にます。若い子羊などの小さな獲物を攻撃する場合、頭蓋骨や脊椎領域を噛むことによって殺害され、組織や骨に大きな損傷を与えます。小さな獲物や若い獲物は完全に持ち去られ、殺害の証拠として血だけが残ることもコヨーテは通常、餌が不足しない限り、皮と大型動物の骨格の大部分を比較的無傷で残しますが、その場合は最大の骨のみを残すことがコヨーテが大きな死骸を大量に食べる場所には、羊毛、皮膚、その他の部分の破片が散在しているのが特徴です。
足跡はコヨーテと犬の捕食を区別する重要な要素です。コヨーテの足跡は飼い犬のものよりも楕円形でコンパクトである傾向があり、爪跡は目立ちにくく、犬の足跡よりも直線に近い傾向がサイトハウンドを除いて、コヨーテと同じ体重のほとんどの犬の歩幅はわずかに短くなります。コヨーテの殺害は、前者の方が下層の組織への損傷が少ないという点でオオカミの殺害と区別することができる。また、コヨーテの糞はオオカミの糞よりも小さい傾向が
コヨーテはドッグフードや獲物として現れるほど小さい動物に引き寄せられることがよくゴミ、ペットフード、場合によっては鳥やリスの餌場などの物品がコヨーテを裏庭に引き寄せます。サウスオレンジカウンティ(カリフォルニア州)の動物緊急治療病院には、コヨーテに襲われた約3~5匹のペットが毎週持ち込まれているが、通常、猫は攻撃を受けても生き残ることができないため、その大半は犬である。カリフォルニア州クレアモント近郊で収集された糞尿の分析により、コヨーテは冬と春の食料源としてペットに大きく依存していることが明らかになった。
南カリフォルニアのある場所では、コヨーテが野良猫のコロニーを食料源として頼り始めました。時間が経つにつれて、コヨーテはほとんどの猫を殺し、その後、猫のコロニーを維持する人々によってコロニーの場所に毎日置かれたキャットフードを食べ続けました。コヨーテは通常、小型の犬を攻撃しますが、例外的にロットワイラーのような大型で強力な犬種さえも攻撃することが知られています。グレイハウンドなど、コヨーテより大きい犬は一般にコヨーテを追い払うことができ、コヨーテを殺すことが知られています。小型犬種は怪我や死亡に見舞われる可能性が高くなります。
狩猟
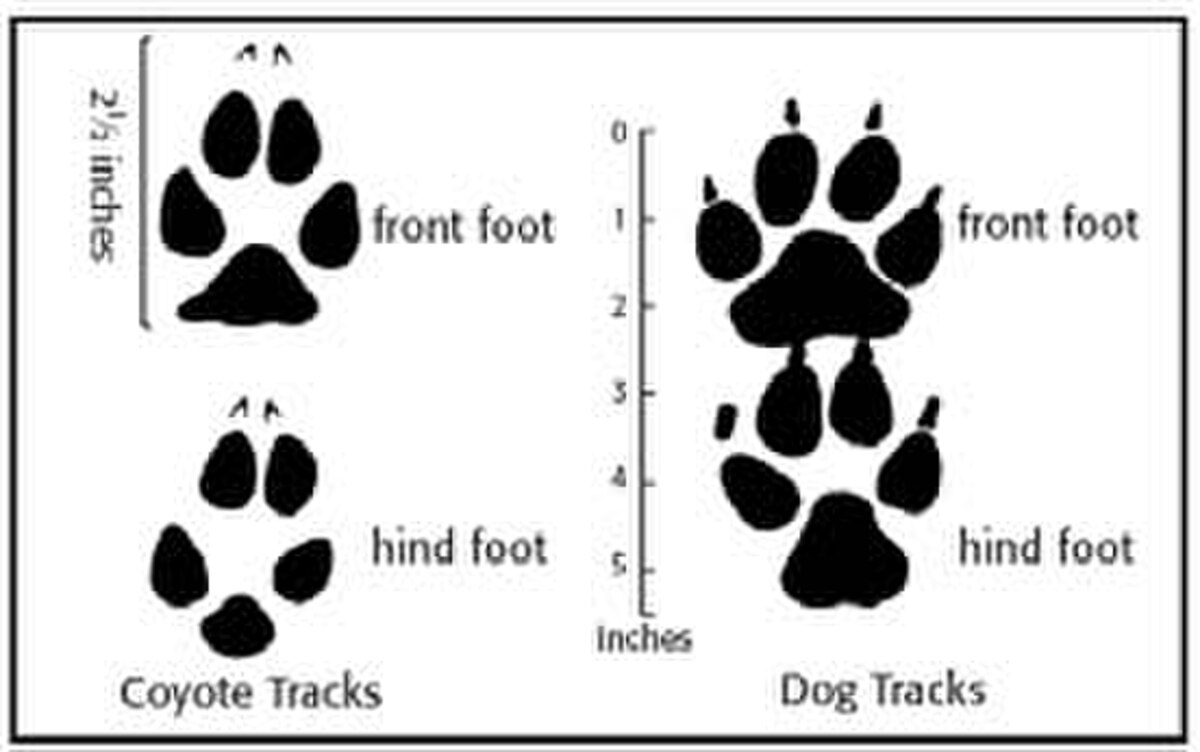
コヨーテの足跡と飼い犬の足跡の比較
コヨーテ狩りは、人間が参加する捕食動物狩りの最も一般的な形式の 1 つです。コヨーテの捕獲に関しては多くの規制がありません。つまり、コヨーテを狩るのに使用できるさまざまな方法が数多く最も一般的な形態は、罠猟、呼び出し猟、猟犬猟です。コヨーテは色盲であるため、グレーと微妙な青の色合いでしか見ることができず、オープンな迷彩や無地のパターンを使用できます。平均的なオスのコヨーテの体重は 8 ~ 20 kg (18 ~ 44 ポンド)、平均的なメスのコヨーテは 7 ~ 18 kg (15 ~ 40 ポンド) であるため、これらの体重の間で性能を発揮できる汎用発射体は .223 レミントンです。発射体はターゲット内で進入後、射出前に膨張するため、最大のエネルギーが供給されます。
コヨーテは軽くて機敏な動物であるため、地形に非常に軽い印象を残すことがよくコヨーテの足跡は長方形で、長さは約 6.35 cm (2.5 インチ)、幅は 5.08 cm (2 インチ) です。前足と後足の両方に4本の爪がコヨーテのセンターパッドは比較的丸い三角形のような形をしています。イエイヌと同様に、コヨーテの前足は後足よりわずかに大きいです。コヨーテの足は飼い犬の足に最も似ています。
毛皮の用途
![]()
カナダ産コヨーテの毛皮
19世紀半ばまでは、コヨーテの毛皮は価値がないと考えられていました。これはビーバーの減少とともに変化し、1860 年までに毛皮を目的としたコヨーテの狩猟はグレート プレーンズのオオカミにとって大きな収入源 (皮あたり75セントから1.50ドル) になりました。コヨーテの毛皮は 1950 年代初期には経済的に非常に重要であり、価格は地域によって異なりますが、1 枚あたり 5 ドルから 25 ドルの範囲でした。コヨーテの毛皮は敷物を作るほど耐久性はないが 、コートやジャケット、スカーフ、マフなどに使用できる。毛皮の大部分は、婦人服のコートの襟や袖などの装飾品の製造に使用されます。コヨーテの毛皮は、イミテーションシルバーフォックスとして黒く染められることも
西部拡大中に、コヨーテは時々わな猟師や山人に食べられることがありました。コヨーテはプレーンズ・インディアンの祝宴に時々登場し、カリフォルニア州サンガブリエルの先住民族はコヨーテの子を食べていました。コヨーテの肉の味はオオカミの味に似ており、茹でると豚肉よりも柔らかくなります。秋に採取されたコヨーテの脂肪は、革に油を塗るために使用されたり、スプレッドとして食べられたりすることが
飼い慣らしやすさ
コヨーテはおそらくコロンビア以前のさまざまな文化によって半家畜化されたと考えられます。19 世紀の作家の中には、グレート プレーンズの原住民の村で飼われているコヨーテについて書いた人もいます。コヨーテは子犬の頃は簡単に飼い慣らされますが、成体になると凶暴になることが純血コヨーテと雑種コヨーテはどちらも遊び好きで飼い主に打ち明けることができるが、疑い深く、見知らぬ人には恥ずかしがり屋であるが、コヨーテは回収や指さしなどの実用的な目的に使用できるほど扱いやすい。記録された。 1945年の夏に捕獲された「ブッチ」という名前の飼いならされたコヨーテは、映画界でのキャリアは短く、鶏小屋を襲撃中に射殺されるまで『スモーキー』(1946年)と『ラムロッド』 (1947年)に出演した。
人気メディアで
ワイリー・E・コヨーテは、短編アニメーション映画の「ルーニー・テューンズ」シリーズと「メリー・メロディーズ」シリーズで顕著に登場します。
アリゾナ州の NHL チームは、この地域に生息するコヨーテの多さに敬意を表して、アリゾナ コヨーテズと名付けられました。
『グッド・ザ・バッド・アンド・アグリー』 (1966年)の有名なウー・ウィー・ウー・ウィー・ワウワウワウの叫び声は、コヨーテの遠吠えからインスピレーションを得たものです。
解説
^ 「ケースドオオカミ」という名前は、歴史的にコヨーテの皮膚がマスクラットの皮膚のようにケース化されていたのに対し、オオカミの皮膚はビーバーのように平らに広げられていたという事実に由来する。
^ サポートする参考文献の完全なセットについては、オオカミの進化#オオカミのようなイヌ科動物の系統樹の注 (a) を参照して
引用
^ テッドフォード、ワン & テイラー 2009、p. 131. ^ Kays, R. (2020) 。「カニス・ラトランス」。IUCN の絶滅危惧種のレッドリスト。2018 : e.T3745A163508579。土井: 10.2305/IUCN.UK.2018-2.RLTS.T3745A163508579.en。2021 年11 月 11 日に取得。
^ ジェームズ、エドウィン; ロング、スティーブン・H. 言って、トーマス。ジョン・アダムズ(1823年)。1819 年と 2020 年に行われたピッツバーグからロッキー山脈への遠征の記録。Vol. 1. ロンドン:ロングマン、ハースト、ピーズ、オーレ、ブラウン。168–174ページ。
^ “ Canis latrans ” . Fossilworks.org 。2021 年12 月 17 日に取得。
^ ベコフ M. (1977)。「カニス・ラトランス」。哺乳類の種(79): 1-9。土井:10.2307/3503817。ISSN 1545-1410。JSTOR 3503817。OCLC 46381503。 ^ ヤング&ジャクソン、1978年、p. 48 ^ ヤング & ジャクソン 1978、63–4 ページ
^ ヤング&ジャクソン、1978年、50–53ページ ^ ヤング&ジャクソン、1978年、p. 247
^ フォックス 1978、p. 105 ^ 「オオカミと土地を共有する」(PDF) . ウィスコンシン州天然資源局。2015年。2016 年6 月 29 日に取得。
^ カルタイノ 2011、p. 16 ^ ヤング&ジャクソン、1978年、p. 59 ^ スティーブン・ヴァンタッセル (2012). 「コヨーテ」。鳥獣被害検査ハンドブック(第 3 版)。ネブラスカ州リンカーン: 野生生物管理コンサルタント。p. 112.ISBN _ 978-0-9668582-5-9。OCLC 794471798。
^ Nowak 1979、p. 14. ^ ホフマイスター、ドナルド F. (2002). イリノイ州の哺乳類。イリノイ大学出版局。33~34ページ。ISBN 978-0-252-07083-9。OCLC 50649299。
^ マッスルマン、ジョセフ 。「コヨーテ」。ルイス&クラークの発見。2013 年 7 月 21 日のオリジナルからアーカイブ。2013 年1 月 15 日に取得。
^ マッスルマン、ジョセフ 。「トーマス・セイ:Canis latrans」。ルイス&クラークの発見。2013 年 7 月 21 日のオリジナルからアーカイブ。2013 年1 月 15 日に取得。
^ クラビジェロ、フランシスコ・ハビエル; チャールズ・カレン(1817年)。メキシコの歴史: スペイン人とメキシコの歴史家、インディアンの写本や古代絵画から収集: スペイン人によるメキシコの征服とともに: メキシコの土地、動物、住民に関する批判的な論文を含む彫刻で描かれています。Vol. 1. フィラデルフィア:トーマス・ドブソン。p. 57. OCLC 13601464。 ^ ブロック、W. (1824)。メキシコでの 6 か月の滞在と旅行: ヌエバ スペインの現状、自然生産物、社会の現状、製造業、貿易、農業、遺物などについての意見が含まれています。: プレートと地図付き。ロンドン: ジョン・マレー、アルベマール・ストリート。119、261ページ。
^ シートン 1909 年、p. 789
^ k l Gier、HT (1974)。「コヨーテ ( Canis latrans ) の生態と行動」。フォックス、MW(編)。野生のイヌ科動物: その体系、行動生態、進化。ニューヨーク:ヴァン・ノストランド・ラインホルト。247–262ページ。ISBN 978-0-442-22430-1。OCLC 1093505。
^ g マーク・ベコフ; ギース、エリック M. (2003)。「コヨーテ」。フェルドハマーでは、ジョージ A. トンプソン、ブルース・C。チャップマン、ジョセフ A. (編)。北アメリカの野生哺乳類: 生物学、管理、および保全(第 2 版)。メリーランド州ボルチモア:ジョンズ・ホプキンス大学出版局。467–470ページ。ISBN
978-0-8018-7416-1。OCLC 51969059。
^ レーナー、フィリップ N. (1978)。「コヨーテコミュニケーション」。ベコフ、M. (編)。コヨーテ: 生物学、行動、管理。ニューヨーク:アカデミックプレス。127–162ページ。ISBN 978-1-930665-42-2。OCLC 52626838。
^ g h ijk ヤング & ジャクソン 1978 、 6–7 ページ ^ カーティス、ES (1928)。チペウィアン。クリーの西部の森。サルシ族。北アメリカのインディアン。Vol. 18. 古典書籍会社。p. 201.ISBN _ 978-0-7426-9818-5。
^ クロフォード、JM (1989)。ココパ辞典。カリフォルニア大学出版局。p. 445.ISBN _ 978-0-520-09749-0。OCLC 20012309。
^ レクレア、N.; カーディナル、G. (1998)。アルバータ州エルダーズクリー語辞典。アルバータ大学。p. 279.ISBN _ 978-0-88864-284-4。OCLC 659111819。
^ マーティン、日本; モールディン、MM (2004)。クリーク/マスコギーの辞典。ネブラスカ大学出版局。p. 153.ISBN _ 978-0-8032-8302-2。OCLC 43561668。
^ アルバート、R.; ショール、DL (1985)。簡潔なホピ語と英語の辞書。ジョン・ベンジャミンズ出版。p. 26.ISBN _ 978-90-272-2015-8。OCLC 777549431。
^ ブライト、ウィリアム; ゲール、スーザン。「コヨーテ」。Karuk の辞書とテキスト。カルク族とカリフォルニア大学バークレー校。2015 年5 月 22 日に取得。
^ リード、FA (2009)。中央アメリカとメキシコ南東部の哺乳類のフィールドガイド。オックスフォード大学出版局。p. 259.ISBN _ 978-0-19-534322-9。OCLC 237402526。
^ 青木春夫 (1994). ネズ・ペルセ辞書。カリフォルニア大学出版局。p. 491.ISBN _ 978-0-520-09763-6。OCLC 463788185。
^ ノインドルフ、A. (1983)。ナバホ語と英語の対訳辞書: Áłchíní Bi Naaltsoostsoh。ニューメキシコ大学出版局。p. 512.ISBN _ 978-0-8263-3825-9。OCLC 57357517。
^ キンテロ、C. (2004)。オーセージ文法。ネブラスカ大学出版局。p. 83.ISBN _ 978-0-8032-3803-9。OCLC 57614396。
^ RP、パークス; ニューサウスウェールズ州プラット (2008)。スキリ・ポーニーの辞書。ネブラスカ大学出版局。p. 119.ISBN _ 978-0-8032-1926-7。OCLC 940905155。
^ デイリー、JP (1989)。トゥンピサ (パナミント) ショショーニ語辞典。カリフォルニア大学出版局。p. 436.ISBN _ 978-0-520-09754-4。OCLC 489876664。
^ ピトキン、H. (1985)。Wintu 辞書。カリフォルニア大学出版局。65、573ページ。ISBN 978-0-520-09613-4。OCLC 12313411。
^ 「コヨーテ」。ユロック辞書: Segep。ユロック言語プロジェクト。カリフォルニア大学バークレー校。2015 年5 月 22 日に取得。
^ “自然史: イヌ科の家族の絆” . アメリカ自然史博物館の雑誌。Vol. 117、いいえ。6. ニューヨーク:アメリカ自然史博物館。2008.p. 22.
^ “ Canis lepophagus ” . 化石工場。2021 年12 月 17 日に取得。
^ ワン & テッドフォード 2008、p. 58. ^ テッドフォード、ワン & テイラー、2009 年、175、180 ページ。
^ ジョンストン 1938、p. 385. ^ Nowak 2003、p. 241. ^ RM ノワク (1978)。「コヨーテと関連するイヌの進化と分類」。ベコフ、M. (編)。コヨーテ: 生物学、行動、管理。ニューヨーク:アカデミックプレス。3–16ページ。ISBN 978-1-930665-42-2。OCLC 52626838。
^ テッドフォード、ワン & テイラー 2009、p. 136. ^ ミーチェン、JA; サミュエルズ、JX (2012)。「巨大動物の絶滅に応じたコヨーテ ( Canis latrans ) の進化」 。米国科学アカデミーの議事録。109 (11): 4191–6。Bibcode : 2012PNAS..109.4191M。土井:10.1073/pnas.1113788109。ISSN 1091-6490。OCLC 475396714。PMC 3306717。PMID 22371581。
^ ミーチェン、JA; ヤノヴィッツ、AC; ジョージア州エイブリー。サドレア、RW (2014)。「氷河期の巨大動物絶滅に応じたコヨーテ ( Canis latrans ) の生態学的変化」 。プロスワン。9 (12): e116041。Bibcode : 2014PLoSO…9k6041M。土井:10.1371/journal.pone.0116041。PMC 4281224。PMID 25551387。 ^ グーレ、GD (1993)。北極、現生、完新世、更新世後期のハイイロオオカミ (Canis lupus) と選択されたイヌ属の間の時間的および地理的頭蓋骨の変異の比較(修士論文)。マニトバ大学ウィニペグ校。1–116ページ。
^ 張、紅海; チェン、レイ (2010)。「ドール Cuon alpinus の完全なミトコンドリア ゲノム: 系統解析とイヌ科内の進化的分岐の年代測定」。分子生物学レポート。38 (3): 1651–60。土井:10.1007/s11033-010-0276-y。PMID 20859694。S2CID 7476436。 ^ フォンホルト、BM; ケーヒル、JA; ファン、Z。グローナウ、I. ロビンソン、J. ポリンジャー、JP; シャピロ、B. ウォール、J. ウェイン、RK (2016)。「全ゲノム配列分析により、北米オオカミの 2 つの固有種はコヨーテとハイイロオオカミの混合であることが示されました。」科学の進歩。2 (7): e1501714。Bibcode : 2016SciA….2E1714V。土井:10.1126/sciadv.1501714。PMC 5919777。PMID 29713682。
^ バージニア州モレル (2016)。「実際にはオオカミではないオオカミをどうやって救うのですか?」科学。353 (6300)。土井:10.1126/science.aag0699。
^ ワン & テッドフォード 2008、p. 。
^ ゴパラクリシュナン、シャム; サマニエゴ・カストゥルイタ、ホセ・A. シンディング、ミッケル・ホルガー S. クデルナ、ルーカスFK; ライコネン、ヤニッケ。ベント州ピーターセン。シケリッツ・ポンテン、トーマス。ラーソン、グレーガー。オーランド、ルドヴィック。マルケス・ボネ、トーマス; ハンセン、アンダース J. ダレン、愛してギルバート、M. トーマス P. (2017)。「オオカミの参照ゲノム配列 (Canis lupus lupus) とイヌ属の集団ゲノミクスへのその意味」。BMC ゲノミクス。18 (1): 495.土井: 10.1186/s12864-017-3883-3。PMC 5492679。PMID 28662691。 ^ シンディング、ミッケル=ホルガー S.; ゴパラクリシャン、シャム。ヴィエイラ、フィリペ G. サマニエゴ・カストゥルイタ、ホセ・A. ラウンドルップ、カトリーヌ。ハイデ・ヨルゲンセン、マッツ・ピーター。モルテン、メルドガード。ベント州ピーターセン。シケリッツ・ポンテン、トーマス。ミケルセン、ヨハン・ブルス。マルカール・ピーターセン、ウルフ; ディーツ、ルーン。ゾンネ、クリスチャン。ダレン、愛してバックマン、ルッツ。ウィグ、オイスタイン。ハンセン、アンダース J. ギルバート、M. トーマス P. (2018)。「北米におけるハイイロオオカミとオオカミに似たイヌ科動物の集団ゲノミクス」。PLOS ジェネティクス。14 (11)。e1007745。土井:10.1371/journal.pgen.1007745。PMC 6231604。PMID 30419012。 ^ ゴパラクリシュナン、シャム; シンディング、ミッケル・ホルガー S. ラモス・マドリガル、ジャズミン。ニーマン、ジョナス。サマニエゴ・カストゥルイタ、ホセ・A. ヴィエイラ、フィリペ G. カロエ、クリスチャン。モンテロ、マルク・デ・マヌエル。クデルナ、ルーカス。セレス、アイトール。ゴンサレス=バサロット、ビクトル・マヌエル。リュウ・ヤン・フー。王国東。マルケス・ボネ、トーマス; ミララブ、シアヴァシュ。フェルナンデス、カルロス。ゴーベール、フィリップ。ケフリ、クラウス・ピーター。バッド、ジェーン。ルーネス、イーライ・ニスペル。ハイデ=ヨルゲンセン、マッツ・ピーター。ベント州ピーターセン。シケリッツ・ポンテン、トーマス。バックマン、ルッツ。ウィグ、オイスタイン。ハンセン、アンダース J. ギルバート、M. トーマス P. (2018)。「種間の遺伝子の流れがイヌ属の進化を形作った」。現在の生物学。28 (21): 3441–3449.e5。土井:10.1016/j.cub.2018.08.041。PMC 6224481。PMID 30344120。 ^ ウォズンクラフト、WC (2005)。「食肉目オーダー」。デラウェア州ウィルソンにて; リーダー、DM (編)。世界の哺乳類種: 分類学的および地理的参照(第 3 版)。ジョンズ・ホプキンス大学出版局。532–628ページ。ISBN
978-0-8018-8221-0。OCLC 62265494。
^ ヤング&ジャクソン、1978年、p. 249 ^ g h i メリアム、スイス (1897)。「新しい形態の記述を伴うコヨーテまたはプレーリーオオカミの改訂」。ワシントン生物学協会の議事録。11:19~33。
^ r s Nowak 1979 、 pp. 9–10
^ エリオット、DG (1904)。中米と西インド諸島の陸と海の哺乳類。Vol. II. イリノイ州シカゴ。467–468ページ。
^ ニューヨーク州ネルソン (1932)。「コヨーテに関するコメント、サルバドール産の新亜種についての説明」。ワシントン生物学協会の議事録。45 : 223–225。2014 年 9 月 7 日のオリジナルからアーカイブ。2014 年9 月 7 日に取得。
^ メンデス=カルバハル、P. (2014)。「哺乳綱、食肉目、イヌ科、イヌ属(セイ、1823年):パナマにおける実際の分布」。チェックリスト。10 (2): 376–379。土井: 10.15560/10.2.376。ISSN 1809-127X。OCLC 828104819 。2015 年1 月 26 日に取得。 ^ スイス、メリアム (1904)。「メキシコ南部からの新種のコヨーテ」。ワシントン生物学協会の議事録。17 : 157. 2014年9月7日時点のオリジナルよりアーカイブ。
^ EA ゴールドマン (1936)。「ホンジュラスからの新しいコヨーテ」。J. ウォッシュ. アカデミア 科学。26:32~34。
^ ヤング&ジャクソン、1978年、p. 263
^ スイス州タウンゼント(1912)。「1911 年にローワー カリフォルニアの『アホウドリ』探検隊によって収集された哺乳類。新種の記載もある。 」アメリカ自然史博物館の紀要。31:117-130。
^ ベイリー、V. (1905)。「テキサスの生物調査」。北アメリカの動物相。25:1-222。土井:10.3996/nafa.25.0001。hdl : 2027/mdp.39015006867405。ISSN 1944-4575。OCLC 273060038。
^ ジャクソン、HHT (1949)。「米国から来た2匹の新しいコヨーテ」。ワシントン生物学協会の議事録。62:31~32。
^ アンダーソン、TM; フォンホルト、BM; シリコーン州キャンディール。ムジアーニ、M. グレコ、C.スターラー、DR; スミス、DW。パドゥカサハスラム、B. ランディ、E。レナード、JA; ブスタマンテ、CD; オストランダー、EA; タン、H. ケンタッキー州ウェイン。バーシュ、GS (2009)。「北米ハイイロオオカミのメラニズムの分子進化史」。科学。323 (5919): 1339 ~ 1343 年。Bibcode : 2009Sci…323.1339A。土井: 10.1126/science.1165448。ISSN 1095-9203。OCLC 34298537。PMC 2903542。PMID 19197024。 ^ ヤング & ジャクソン 1978、121–24 ページ ^ カルタイノ 2011、61–3 ページ
^ フォックス 1978、p. 136 ^ カール、ジマー (2013 年 1 月 21 日)。「スノーコヨーテとスピリットベア」。ナショナル ジオグラフィック誌。2016 年7 月 1 日に取得。
^ ウェイ、JG; ラトリッジ、L. ウィールドン、T. BN ホワイト (2010)。「マサチューセッツ州東部の東部「コヨーテ」の遺伝的特徴付け」(PDF)。北東部の博物学者。17 (2): 189–204。土井:10.1656/045.017.0202。ISSN 1938-5307。JSTOR 40664873。OCLC 46381506。S2CID 135542。2011 年 1 月 7 日のオリジナルからアーカイブ(PDF) 。
^ やあ、JG (2007). 「北アメリカ東部と西部におけるイヌ ラトランス(コヨーテ)の体重の比較」 (PDF)。北東部の博物学者。14 (1): 111-24。土井:10.1656/1092-6194(2007)142.0.CO;2。ISSN 1938-5307。JSTOR 4499900。OCLC 46381506。S2CID 85288738。2008 年 10 月 6 日のオリジナルからアーカイブ(PDF) 。
^ ベコフ、M. (1978)。「コヨーテとイースタンコヨーテの行動発達」。コヨーテ: 生物学、行動、管理。ニューヨーク:アカデミックプレス。97–127ページ。ISBN 978-1-930665-42-2。OCLC 52626838。
^ ヘンリー・ヒルトン (1978). 「イースタンコヨーテの系統学と生態学」。ベコフ、M. (編)。コヨーテ: 生物学、行動、管理。ニューヨーク:アカデミックプレス。210–28ページ。ISBN 978-1-930665-42-2。OCLC 52626838。
^ フォンホルト、BM; ポリンジャー、JP; アール、DA; ノウルズ、JC。アーカンソー州ボイコ。パーカー、H. ゲフィン、E.パイロット、M。Jedrzejewski、W. Jedrzejewska、B. シドロヴィッチ、V。グレコ、C.ランディ、E。ムジアーニ、M. ケイズ、R. ブスタマンテ、CD; オストランダー、EA; ノベンバー、J. ウェイン、RK (2011)。「謎めいたオオカミに似たイヌ科動物の進化史に関するゲノムワイドな視点」。ゲノム研究。21 (8): 1294–1305。土井:10.1101/gr.116301.110。ISSN 1549-5469。OCLC 37589079。PMC 3149496。PMID 21566151。
^ フォックス、MW (1974)。「イヌ科動物の社会行動の進化」。野生のイヌ科動物: その体系、行動生態学、および進化。ニューヨーク:ヴァン・ノストランド・ラインホルト。429–60ページ。ISBN 978-0-442-22430-1。OCLC 1093505。
^ マーク・ベコフ; ダイヤモンド、ジュディ (1976)。「コヨーテの交尾前および交尾行動」。哺乳類ジャーナル。57 (2): 372–375。土井:10.2307/1379696。ISSN 0022-2372。JSTOR 1379696。OCLC 1800234。 ^ デブラ・A・カールソン; ギース、エリック M. (2008)。「コヨーテ (Canis latrans) の生殖生物学: 交尾行動、生殖ホルモン、および膣細胞学の統合」。哺乳類ジャーナル。89 (3): 654–664。土井:10.1644/06-mamm-a-436r1.1。PMC 7108653。PMID 32287378。
^ ギース、エリック M.; ラフ、ロバート L. (1997)。「コヨーテ、イヌの匂いマーキング:社会的および生態学的要因の影響」。動物の行動。54 (5): 1155–1166。CiteSeerX 10.1.1.540.1024。土井:10.1006/anbe.1997.0561。PMID 9398369。S2CID 33603362。 ^ メカ、DL (2003)。ミネソタのオオカミ: ハートランドの遠吠え。ヴォイジャープレス。p. 75.ISBN _ 978-0-89658-509-6。OCLC 43694482。
^ カリフォルニア州ヘネシー; ドゥバック、J. ゲールト、SD (2012)。「都市部のコヨーテ間の長期的なペア結合と一夫一婦制の遺伝的証拠 ( Canis latrans )」。哺乳類ジャーナル。93 (3): 732–742。土井: 10.1644/11-MAMM-A-184.1。ISSN 1545-1542。OCLC 39098574。 ^ フォックス 1978、p. 33 ^ ムロット、クリス (1998)。「ラマーバレーのコヨーテ: イエローストーンでは、マスターアダプターがオオカミへの対処法を学びます。」科学ニュース。153 (5): 76-78。土井:10.2307/4010114。JSTOR 4010114。
^ ウェルズ、マイケル C.、マーク ベコフ。「コヨーテ Canis latrans の香りマーキングの観察研究」(1981年)。
^ ヤング&ジャクソン、1978年、82–90ページ ^ アサ、CS; Mech、D. (1995)。「オオカミの感覚器官と生活史におけるそれらの重要性の概説」。LD州カービンにて。フリッツ、SH; ザイプ、DR (編)。変化する世界におけるオオカミの生態と保護。アルバータ州エドモントン: アルバータ大学。287–291ページ。ISBN 978-0-919058-92-7。OCLC 35162905。
^ エステルホルム、H. (1964)。「キツネ ( Vulpes vulpes L. )の摂食行動における距離受信の重要性」。アクタ ズーロジカ フェニカ。106 : 1-31.
^ ウェルズ、MC (1978)。「コヨーテは捕食中に感覚を感じる – コヨーテの相対的な利用に対する環境の影響」。行動プロセス。3 (2): 149-158。土井:10.1016/0376-6357(78)90041-4。PMID 24924653。S2CID 22692213。 ^ ウェルズ、MC; レーナー、ペンシルバニア州 (1978)。「コヨーテの捕食行動における距離感の相対的な重要性」。動物の行動。26 : 251–258。土井:10.1016/0003-3472(78)90025-8。S2CID 53204333。 ^ ヤング&ジャクソン、1978年、91–92ページ ^ ヤング&ジャクソン、1978年、p. 97 ^ ヤング&ジャクソン、1978年、p. 98 ^ ジェイコブス、ジュリア (2018 年 6 月 15 日)。「哺乳類は人間を避けるために夜行性になる」ニューヨークタイムズ紙。2022 年 1 月 1 日のオリジナルからアーカイブ。2018 年6 月 21 日に取得。
^ ゲイナー (2021年6月15日). 「野生動物の夜行性に対する人間の妨害の影響」。科学。360 (6394): 1232–1235。土井:10.1126/science.aar7121。PMID 29903973。S2CID 49212187 。2021 年11 月 14 日に取得。 ^ 米国魚類野生生物局 (2016)。「発見!一緒に狩りをするコヨーテとアナグマ」
^ ヤング&ジャクソン 1978、pp. 93–96 ^ “なぜ私の庭にコヨーテがいるのですか?餌の餌とその他の答え” . 米国人道協会。2020 年5 月 7 日に取得。
^ シルバー、H.; シルバー、ウェストバージニア州 (1969)。「ニューイングランド北部のコヨーテに似たイヌ科の成長と行動、およびイヌ科の雑種に関する観察」。野生生物協会、野生生物モノグラフ。17 (17):24-25。ISSN 1938-5455。JSTOR 3830473。OCLC 60618095。 ^ フォックス 1978、134–135 ページ ^ ビー、ジェームス (1981). カンザス州の哺乳類。カンザス大学。p. 165.
^ マイケル・A・メレス; オクラホマ自然史博物館 (オクラホマ州ノーマン) (1999)。砂漠の百科事典。オクラホマ大学出版局。137–8ページ。ISBN 978-0-8061-3146-7。
^ フランコウィアク、グレゴリー A.; ペルディカス、マルロ。スミス、グレゴリー A. (2019 年 12 月 30 日)。「オハイオ州カヤホガ渓谷の都市化する風景におけるコヨーテの空間生態学」。プロスワン。14 (12): e0227028。ビブコード: 2019PLoSO..1427028F。土井:10.1371/journal.pone.0227028。ISSN 1932-6203。PMC 6936805。PMID 31887177。 ^ ワシントン州サンダース「アディロンダック生態センター: コヨーテ」。ニューヨーク州環境科学林業大学。
^ ロバート・L・ホール; シャープ、ヘンリー S. (1978)。オオカミと人間: 並行進化。ニューヨーク:アカデミックプレス。p. 156.ISBN _ 978-0-12-319250-9。OCLC 3607816。
^ サウスカロライナ州ミンタ; カンザス州ミンタ。ロット、DF (1992)。「アナグマ (Taxidea Taxus) とコヨーテ (Canis latrans) の間の狩猟関連性」。哺乳類ジャーナル。73 (4): 814–820。土井:10.2307/1382201。JSTOR 1382201。 ^ バーテル、RA; ノルトン、FF (2005)。「ユタ州カーリューバレーにおける獲物の量の変動に対するコヨーテ、イヌ・ラトランスの機能的な摂食反応、1977年から1993年」。カナダ動物学ジャーナル。83 (4): 569–578。土井:10.1139/z05-039。
^ よろしくお願いします、TL; ホディチェック、B. トーマス、HH (1981)。「オクラホマ州のコヨーテ (Canis latrans) の食べ物」。南西部の博物学者。26 (1): 67-69。土井:10.2307/3671333。JSTOR 3671333。 ^ トカール、エリック。「イヌ・ラトランス」。動物多様性ウェブ。2023 年2 月 16 日に取得。
^ ブランディジ、GC (1993)。ニューヨーク州アディロンダック中央部におけるイースタンコヨーテ ( Canis latrans var.) の捕食生態(PhD)。ニューヨーク州立大学、環境科学林業カレッジ、シラキュース。
^ ボイヤー、RH (1949)。「セコイア国立公園でマウンテンコヨーテが一歳のツキノワグマを殺す」哺乳類ジャーナル。30 : 75.土井: 10.1093/jmammal/30.1.75。ISSN 1545-1542。OCLC 39098574。 ^ さすが、JG; ホートン、J. (2004)。「コヨーテがタテゴトアザラシを殺す」(PDF)。カニドニュース。7 (1)。ISSN 1545-1542。OCLC 39098574。2006 年 5 月 13 日のオリジナル(PDF)からアーカイブ。
^ クラウバー、ローレンス・モンロー (1997). ガラガラヘビ:その習性、生活史、人類への影響。Vol. 1(第2版)。カリフォルニア州バークレー:カリフォルニア大学出版局。1072–1074ページ。ISBN 978-0-520-21056-1。OCLC 39523012。
^ スペリー、CC (1939)。「ペグレッグコヨーテの食性」。哺乳類ジャーナル。20 (2): 190–194。土井:10.2307/1374376。JSTOR 1374376。 ^ マクラッケン、JG; ワシントン州ウレシュ (1984)。「サウスダコタ州ブラックヒルズのコヨーテ料理」。野生生物管理ジャーナル。48 (4): 1420–1423。土井:10.2307/3801809。JSTOR 3801809。 ^ Smith, JW (1988)、「ミズーリ州の実験的トランペッター・スワン修復プログラムの状況」、D. Compton (編)、In Proc. および第10回トランペッター白鳥協会会議の論文 、ミネソタ州メープル プレイン: トランペッター白鳥協会、100–103 ページ
^ フロリダ州バネル; ダンバー、D. コザ、L. ライダー、G. (1981)。「ブリティッシュコロンビア州のモモイロペリカンの生産性と数に対する撹乱の影響:観察とモデル」。コロニアル水鳥。4:2~11。土井:10.2307/1521105。JSTOR 1521105。
^ ギース、EM; オジサマ州ロンスタッド。ミットン、WR (1988)。「コロラド州南東部におけるコヨーテの群れのサイズと食事の関係」。野生生物管理ジャーナル。52 (4): 647–653。土井:10.2307/3800924。JSTOR 3800924。 ^ WD、ボーエン (1978)。獲物のサイズに関連したコヨーテの社会的組織(PhD.)。ブリティッシュコロンビア大学。
^ アルジョ、WM; プレッチャー、DH。リーム、RR (2002)。「モンタナ州北西部におけるオオカミとコヨーテの食生活の重複」。哺乳類ジャーナル。83 (3): 754–766。土井:10.1644/1545-1542(2002)083<0754_dobwac>2.0.co;2。
^ ギース、EM; グローテ、S. (1995)。「ワイオミング州イエローストーン国立公園における冬季のシカとヘラジカに対するコヨーテの捕食の分析」。アメリカのミッドランド博物学者。133 (1): 36–43。土井:10.2307/2426345。JSTOR 2426345。 ^ HA州ウィットロー; バラード、WB。サビーヌ、DL; ヤング、SJ。ジェンキンス、RA; フォーブス、ジョージア州 (1998)。「ニューブランズウィック州における成体のオジロジカの生存率と原因別死亡率」。野生生物管理ジャーナル。62 (4): 1335–1341。土井:10.2307/3801999。JSTOR 3801999。 ^ ブランズ、EH (1970)。「イヌワシとコヨーテのプロングホーンアンテロープに対する冬の捕食」。できる。フィールドナット。84 : 301–304。
^ ボーグス、EK; アンドリュース、RD; ビショップ、RA (1978)。「アイオワ州におけるコヨーテと犬による家畜の損失」。野生生物管理ジャーナル。42 (2): 362–372。土井:10.2307/3800272。JSTOR 3800272。 ^ Connolly, G. (1992)、家畜およびその他の資源に対するコヨーテの被害。ボーア、AH、イースタンコヨーテの生態と管理、ニューブランズウィック州: ニューブランズウィック大学
^ リード、レイチェルEB; ギフォード=ゴンザレス、ダイアン。コッホ、ポール L (2018)。「カリフォルニア沿岸におけるコヨーテ (Canis latrans) の海洋資源の利用: 最近の祖先と比較した新しい行動」。完新世。28 (11): 1781–1790。Bibcode : 2018Holoc..28.1781R。土井: 10.1177/0959683618788714。
^ コーディ、ヒュー (2013). 北アメリカ: 1 つの大陸にある世界。フィラデルフィア:ランニングプレス。ISBN 978-0-7624-4843-2。OCLC 808413615。
^ バーガー、KM; ギース、EM (2007)。「オオカミとの干渉競争がコヨーテの分布と個体数を制限するのでしょうか?」。動物生態学ジャーナル。76 (6): 1075–1085。土井: 10.1111/j.1365-2656.2007.01287.x。PMID 17922704。 ^ ステイン、HJ (1974)。「イヌ科の分布と分類」。フォックス、MW(編)。野生のイヌ科動物: その体系、行動生態学、および進化。ニューヨーク:ヴァン・ノストランド・ラインホルト。3–26ページ。ISBN 978-0-442-22430-1。OCLC 1093505。
^ ホーノッカー、M. ネグリ、S. (2009)。クーガー: エコロジーと保全。シカゴ大学出版局。p. 170.ISBN _ 978-0-226-35347-0。OCLC 609634655。
^ パロマレス、F.; カロ、TM (1999)。「哺乳類肉食動物間の種間殺害」(PDF)。アメリカの博物学者。153 (5): 492–508。土井:10.1086/303189。hdl : 10261/51387。PMID 29578790。S2CID 4343007。
^ BB アッカーマン、FG リンゼイ、TP ヘムカー (1984)。ユタ州南部のクーガーの食習慣。野生生物管理ジャーナル、147–155。
^ エルナンデス・セント・マルティン、AD; ローザス・ロザス、オクラホマ州。パラシオ=ヌニェス、J. ルイジアナ州タランゴ・アランブラ。クレメンテ・サンチェス、F. アラバマ州ホーゲシュタイン (2015)。「メキシコ北東部の保護区とそれに隣接する断片的な風景におけるジャガーとピューマの食性」。自然地域ジャーナル。35 (2): 308–317。土井: 10.3375/043.035.0213。S2CID 86622145。 ^ ビール、北米哺乳類百科事典: 北アメリカ哺乳類の必須ガイド。サンダーベイ出版 (2004)、
ISBN 978-1-59223-191-1。 ^ CAW、グギスバーグ (1972)。ワニ: その自然史、民俗、および保護。p. 195.ISBN _ 978-0-7153-5272-4。
^ オドナヒュー、M.、ブーティン、S.、マレー、DL、クレブス、CJ、ホーファー、EJ、ブライテンモーザー、U.、ブライテンモーザー=ヴュルステン、C.、ズレタ、G.、ドイル、C. & ナムズ、VO (2001)。コヨーテとオオヤマネコ。北方林の生態系ダイナミクス: Kluane プロジェクト。オックスフォード大学出版局、ニューヨーク、ニューヨーク、米国、276–323。
^ メイソン、JR (2000)。「イヌワシが大人のオスのコヨーテを攻撃して殺害」(PDF)。猛禽研究ジャーナル。34 (3): 244–245。2008 年 9 月 22 日のオリジナルからアーカイブ(PDF) 。
^ アレン、ML; エルブロッホ、LM; ウィルマーズ、CC; ウィットマー、ヒューストン (2015)。「スカベンジャーによる死肉の獲得に対する大型肉食動物の比較影響」。アメリカの博物学者。185 (6): 822–833。土井:10.1086/681004。hdl : 2142/101871。PMID 25996866。S2CID 206003003。 ^ グリーン、GI、マットソン、DJ、およびピーク、JM (1997)。イエローストーン国立公園で、ハイイログマが有蹄類の死骸を食べる春。野生生物管理ジャーナル、1040-1055。
^ ウィルマーズ、CC; スターラー、DR; ロードアイランド州クラブツリー。スミス、DW。ゲッツ、W.M. (2003)。「資源の分散と消費者の優位性:米国イエローストーン都市圏でのオオカミとハンターの死骸のあさり」エコロジーレター。6 (11): 996–1003。土井: 10.1046/j.1461-0248.2003.00522.x。S2CID 55961335。 ^ MG、ホーノッカー; ハッシュ、HS (1981)。「モンタナ州北西部のクズリの生態」。カナダ動物学ジャーナル。59 (7): 1286–1301。土井:10.1139/z81-181。
^ WD、ボーエン (1980)。「有蹄類の死骸におけるコヨーテとイヌワシの相互作用」。J.哺乳類。61 (2): 376–377。土井:10.2307/1380075。JSTOR 1380075。 ^ ヨルドハイム、S (1980)。「イーグル・コヨーテ事件」。ブルージェイ。38:47~48。土井:10.29173/bluejay4577。
^ ジョン、TS; ケンタッキー州エヴェラット。アンドレセン・エヴェラット、LM (2009)。「カナダ北西部におけるイヌワシ (Aquila chrysaetos) によるコヨーテ (Canis latrans) の盗虫寄生」。北西部の博物学者。90 (1): 53-55。土井: 10.1898/1051-1733-90.1.53。S2CID 86356027。 ^ マサチューセッツ州マッコロー; トッド、CS; オーウェン RB ジュニア (1994)。「メイン州の越冬中のハクトウワシに対する追加給餌プログラム」。野生動物協会の会報。22 (2): 147–154。土井:10.1016/0006-3207(96)83230-7。JSTOR 3783240。 ^ 軍曹、アラン B. アレン、スティーブン H. (1989)。「観察されたコヨーテとアカギツネの間の相互作用」。哺乳類ジャーナル。70 (3): 631–633。土井:10.2307/1381437。ISSN 1545-1542。JSTOR 1381437。OCLC 39098574。
^ フェドリアーニ、JM; フラー、TK; RM州ソーバジョット。EC 州ヨーク (2000 年)。「3 つの同所性肉食動物間の競争とギルド内捕食」(PDF)。生態学。125 (2): 258–270。Bibcode : 2000Oecol.125..258F。土井:10.1007/s004420000448。hdl : 10261/54628。ISSN 1432-1939。OCLC 76327396。PMID 24595837。S2CID 24289407。2011 年 10 月 6 日のオリジナル(PDF)からアーカイブ。
^ リトヴァイティス、JA; DJハリソン(1989年)。「コヨーテの個体数増加期におけるボブキャットとコヨーテのニッチな関係」。カナダ動物学ジャーナル。67 (5): 1180–1188。土井:10.1139/z89-170。
^ バネル、ケビン D.; フリンダース、ジェラン・T. ウォルフ、マイケル L. (2006)。「西部西部におけるオオヤマネコの保護に対するコヨーテとスノーモービルの潜在的な影響」。野生動物協会の会報。34 (3): 828–838。土井:10.2193/0091-7648(2006)342.0.CO;2。ISSN 1938-5463。JSTOR 3784713。OCLC 60353682。S2CID 85901240。 ^ ギプソン、PS; JF カムラー (2002)。「コヨーテに殺されたボブキャット」。南西部の博物学者。47 (3): 511–513。土井:10.2307/3672519。ISSN 0038-4909。JSTOR 3672519。OCLC 525604174。 ^ アンダーソン、EM (1986)。コロラド州南東部における資源利用に関するボブキャットの行動生態学。論文、コロラド州立大学、フォートコリンズ、米国。
^ デラウェア州ジャクソン (1986)。コロラド州中東部のボブキャットの生態。論文、コロラド州立大学、フォートコリンズ、米国。
^ デラウェア州トウェイル (1986)。針葉樹林におけるボブキャットとコヨーテによる資源分割。論文、オレゴン州立大学、コーバリス、米国 ^ ニック、ST (1990)。「アイダホ州南東部における搾取と獲物の減少に関するボブキャットの生態」。野生動物モノグラフ。108 (108): 1 ~ 42。JSTOR 3830671。
^ リップル、WJ; ワイジング、AJ。ベシュタ、RL; サウスウェールズ州バスカーク (2011)。「オオカミの回復はオオヤマネコの回復に役立ちますか?」(PDF)。野生動物協会の会報。35 (4): 514–518。土井:10.1002/wsb.59。2014 年 4 月 16 日のオリジナルからアーカイブ(PDF) 。
^ オドナヒュー、M.; ホーファー、EJ。ドイル、フロリダ州 (1995)。「プレデター対プレデター」。自然史。104:6-9。
^ ロックウッド、ラリー L. (2015). 人口生態学の入門。イギリス、チチェスター:ジョン・ワイリー・アンド・サンズ。p. 273.ISBN _ 978-1-118-94755-5。OCLC 899267730。
^ サウスウェールズ州ブッシュカーク; ルッジェーロ、LF; クレブス、CJ (2000)。「生息地の分断と種間競争:オオヤマネコの保全への影響」(PDF)。LF、ルッジェーロにて。オーブリー、KB; サウスウェールズ州バスカーク。ケーラーGM。クレブス、CJ; カンザス州マッケルベイ。スクワイアズ、JR (編)。米国におけるオオヤマネコの生態と保護。デンバー:コロラド大学出版局。91-92ページ。2007 年 2 月 26 日のオリジナルからアーカイブ(PDF) 。
^ ホーディ、ジェームス・W; ケイズ、ローランド (2018)。「北および中央アメリカにわたるコヨーテ (Canis latrans) の拡大をマッピングする」。ズーキー(759): 81–97。土井:10.3897/zookeys.759.15149。PMC 5974007。PMID 29861647。 ^ “コヨーテは北アメリカを征服した。今は南へ向かっている” . ニューヨークタイムズ紙。2018 年 5 月 24 日。ISSN 0362-4331。2018年5月24日のオリジナルからアーカイブ。2018 年5 月 25 日に取得。
^ イダルゴ=ミハルト、MG (2004)。「メキシコと中央アメリカにおけるコヨーテ ( Canis latrans ) の歴史と現在の分布」 。生物地理学ジャーナル。31 (12): 2025 ~ 2038 年。土井:10.1111/j.1365-2699.2004.01163.x。S2CID 56048806。2017 年 5 月 25 日のオリジナルからアーカイブ。2015 年1 月 25 日に取得。 ^ デラロサ、CL; ノッケ、CC (2010)。「肉食動物の進化:中央アメリカと南北の大移動」。中央アメリカの肉食獣へのガイド: 自然史、生態学、および保護。テキサス大学出版局。ISBN 978-0-292-78951-7。
^ ヘッペンハイマー、エリザベス; コシオ、ダニエラ S. クリスティン・E・ブレゼスキー; コーディル、ダニー。ヴァン・ホワイ、カイル。チェンバレン、マイケル・J。ヒントン、ジョセフ W. フォンホルト、ブリジット (2017)。「人口動態の歴史は、最近拡大したコヨーテ (Canis latrans) 個体群における遺伝的多様性の空間パターンに影響を与えています。 」遺伝。120 (3): 183–195。土井:10.1038/s41437-017-0014-5。PMC 5836586。PMID 29269931。 ^ ヤング&ジャクソン、1978年、107–114ページ ^ ジャイア、HT; SM、クラッケンバーグ。RJ マーラー (1978)。「コヨーテの寄生虫と病気」。ベコフ、M. (編)。コヨーテ: 生物学、行動、管理。ニューヨーク:アカデミックプレス。37–71ページ。ISBN 978-1-930665-42-2。OCLC 52626838。
^ チャイ、JY; ダーウィン、マレル K. AJ リンベリー (2005)。「魚媒介性人獣共通感染症:現状と問題点」。寄生虫学の国際ジャーナル。35 (11–12): 1233–1254。土井:10.1016/j.ijpara.2005.07.013。PMID 16143336。 ^ ワッツ、LS (2006)。アメリカ民間伝承の百科事典。インフォベース出版。93-94ページ。ISBN 978-1-4381-2979-2。OCLC 465438817。
^ ハリス、M. (1979)。文化的唯物論: 文化科学のための闘争。ニューヨーク: AltaMira Press。200–1ページ。ISBN 978-0-7591-0135-7。OCLC 47100657。
^ アンガス・K・ガレスピー; ジェイ・メクリング (1987)。シンボルとストーリーにおけるアメリカの野生動物。テネシー大学出版局。225–230ページ 。ISBN 978-0-87049-522-9。OCLC 14165533。
^ やあ、JG (2012). 「オオカミが好きでコヨーテが嫌い? イヌ科動物愛好家にとっての難問」(PDF)。インターナショナルウルフ。22 (4):8-11。2012 年 12 月 24 日のオリジナル(PDF)からアーカイブ。
^ ペンシルバニア州リンチ; ロバーツ、J. (2010)。ネイティブ アメリカンの神話 A から Z まで。インフォベース出版。p. 27.ISBN _ 978-1-4381-3311-9。OCLC 720592939。
^ シュワルツ、M. (1998)。初期アメリカにおける犬の歴史。エール大学出版局。146–149ページ。
ISBN 978-0-300-07519-9。 ^ メイン州ミラー; KA タウブ (1993)。古代メキシコとマヤの神々とシンボル: メソアメリカ宗教の図解辞典。テムズ川とハドソン川。p. 92.ISBN 978-0-500-05068-2。OCLC 27667317。
^ オリヴィエ、G. (2003)。アステカ神の嘲笑と変身: テスカトリポカ、「喫煙する鏡の主」。コロラド大学出版局。p. 32.ISBN 978-0-87081-745-8。OCLC 52334747。
^ ケリー、DH (1955)。「ケツァルコアトルと彼のコヨーテの起源」。エル・メキシコ・アンティグオ。8 : 397–416。
^ 「コヨーテの襲撃: 増大する郊外問題」(PDF)。2004 年 3 月。2007年 9 月 26 日のオリジナル(PDF)からアーカイブ。2007 年8 月 19 日に取得。
^ アタック・イン・ザ・ワイルド: コヨーテ・ミステリー(ドキュメンタリー)。ナショナル ジオグラフィック チャンネル。2009 年 10 月 27 日。2016 年11 月 5 日に取得。
^ デラモーレ、クリスティーン 。「シティスリンカーズ」。スミソニアン博物館。2013 年 12 月 20 日のオリジナルからアーカイブ。2012 年6 月 14 日に取得。
^ ベイカー、レックス O. (2007)。「南カリフォルニアにおける人間やペットへの攻撃を防止または軽減するために実施され、成功した都市部コヨーテ管理プログラムのレビュー」。第 12 回野生動物被害管理会議の議事録。382–392ページ。
^ ケイズ、R.; カーティス、A. JJ カーヒマン (2009)。「オオカミとの交雑による北東産コヨーテの急速な適応進化」。生物学の手紙。6 (1): 89-93。土井:10.1098/rsbl.2009.0575。PMC 2817252。PMID 19776058。 ^ “”コヨーテの捕食 – 説明”” . 2007 年 8 月 6 日のオリジナルからアーカイブ。2007 年8 月 19 日に取得。
^ 羊とヤギの死亡事故(PDF) (レポート)。国家農業統計局。2005 年 5 月 6 日。2019年 1 月 22 日のオリジナルからアーカイブ(PDF) 。2020 年11 月 27 日に取得。
^ 羊とヤギの死亡事故(PDF) (レポート)。国家農業統計局。2010 年 5 月 27 日。2019年 6 月 9 日のオリジナルからアーカイブ(PDF) 。2020 年11 月 27 日に取得。
^ 「米国における羊と子羊の捕食者と非捕食者の死亡数、2015年」(PDF) . 米国農務省。2015. 2016 年 4 月 3 日のオリジナルからアーカイブ(PDF) 。2016 年7 月 1 日に取得。
^ “羊と子羊の目録” . 米国農務省。2010 年2 月 1 日に取得。
^ “狡猾なコヨーテを制御する? 簡単な答えはまだない” . NBCニュース。2009 年 12 月 7 日。2013 年9 月 14 日に取得。
^ コリンジ、マーク; ティム、RM; マドン、MB (2008)。「アイダホ州における個々のオオカミ、ツキノワグマ、ピューマ、コヨーテによる家畜捕食の相対的リスク」。脊椎動物害虫会議議事録: 129–133。
^ ユタ州の捕食者制御プログラムの概要、2016 年 7 月 1 日から 2017 年 6 月 30 日までのプログラム活動およびデータ(PDF) (レポート)。ユタ州野生生物資源局。2017年。
^ 「家畜保護犬」(PDF) . 野生動物サービス。2010 年 10 月。2016 年 9 月 9 日のオリジナル(PDF)からアーカイブ。2016 年7 月 3 日に取得。
^ “家畜番犬に関するファクトシート” . 米国農務省動植物衛生検査サービス。2012 年 3 月 24 日のオリジナルからアーカイブ。2012 年4 月 3 日に取得。
^ グランディン、寺院 (2015 年 2 月 26 日)。「専門家らは、適切に行われた牧畜は環境と野生動物の生息地を改善すると述べています。 」牛肉雑誌。2017 年12 月 30 日に取得。
^ “”繁殖牛、繁殖未経産牛、フィーダー子牛”” . Rhino’s ビーフ & 農場で育てられたオーストラリア人。2016 年 10 月 4 日。2017 年12 月 30 日に取得。
^ “オオカミの略奪に関する牧場主向けガイド” . モンタナ州立大学。2006年。2013年4月9日のオリジナルからアーカイブ。2016 年7 月 1 日に取得。 CS1 メイン: URL が不適切です (リンク) ^ ローリンズ、デール。「コヨーテへの対処: 家畜の損失を最小限に抑えるための管理代替案」(PDF)。テキサス農業普及サービス。ページ 4–7 。2016 年11 月 5 日に取得。
^ ハーデスティ、グレッグ (2005 年 5 月 5 日)。「コヨーテにとってペットは獲物です。 」グレッグ・ハーデスティ オレンジ郡登録簿。2007 年 7 月 15 日のオリジナルからアーカイブ。
^ “ウェイマスでコヨーテが襲撃、犬を殺害” . WHDH-TV – ニューイングランドニュース。2007 年 5 月 14 日 ^ マクール、ジュリエット (2010). 「コヨーテ vs. グレイハウンド: 戦線は描かれる」。ニューヨークタイムズ。2022 年 1 月 1 日のオリジナルからアーカイブ。2016 年7 月 3 日に取得。
^ “コヨーテの国: コヨーテ狩りの短期集中コース” . アウトドアライフ。2019年1月23日。2019 年12 月 22 日に取得。
^ “コヨーテの国: コヨーテ狩りの短期集中コース” . アウトドアライフ。2019年1月23日。2020 年5 月 7 日に取得。
^ “”コヨーテの足跡を識別する方法”” . ワイルドライフ ランド トラスト。2020 年5 月 10 日に取得。
^ ヤング&ジャクソン、1978年、115–116ページ ^ シートン 1909、p. 816 ^ ヤング&ジャクソン、1978年、119–21ページ
^ ヤング & ジャクソン 1978、64–9 ページ ^ JW シュルツ (1962)。ブラックフィートとバッファロー:インディアンの人生の記憶。オクラホマ大学出版局。141–3ページ。ISBN 978-0-8061-1700-3。OCLC 248716。
^ エッター、J. (1998 年 2 月 15 日)。「コヨーテはダーラムマンにとって最高の鳥犬として溶け込んでいる」オクラホマン。2016 年7 月 1 日に取得。
^ マーク、エドワーズ (2007 年 4 月 1 日)。「善良な人、勇敢な人、そして聡明な人」。タイムズ。ロンドン。2011 年 5 月 17 日のオリジナルからアーカイブ。2007 年5 月 26 日に取得。
一般的な情報源と引用された情報源
カルタイノ、キャロル (2011)。コヨーテに関する神話と真実: アメリカで最も誤解されている捕食者について知っておくべきこと。Readhowyouwant.com。ISBN 978-1-4587-2668-1。OCLC 876517032。
ミネソタ州フォックス (1978)。犬:その飼いならしと行動。ガーランドSTPMプレス。ISBN 978-0-8240-9858-2。OCLC 3223381。
ジョンストン、CS (1938)。「シタキャニオンの脊椎動物のタイプの産地と祖先コヨーテの記述に関する予備報告」。アメリカ科学ジャーナル。5. 35 (209): 383–390。ビブコード: 1938AmJS…35..383J。土井:10.2475/ajs.s5-35.209.383。
RM ノワク (1979)。「最近の人口の歴史と統計分析」。Wiley、EO (編)。北アメリカ第四紀イヌ科。Vol. 6. カンザス州ローレンス: カンザス大学印刷サービス。ISBN 0-89338-007-5。
RM ノワク (2003)。「オオカミの進化と分類学」。『Mech』では、L. デビッド。ボイタニ、ルイージ (編)。オオカミ: 行動、生態学、保全。シカゴ大学出版局。239–258ページ。ISBN 978-0-226-51696-7。
ET、シートン(1909年)。北方動物の生活史 : マニトバ州の哺乳類の説明。ニューヨーク: スクリブナー。
テッドフォード、リチャード・H. 王暁明。テイラー、ベリル E. (2009)。「北米化石イヌ科(食肉目:イヌ科)の系統体系」 (PDF)。アメリカ自然史博物館の紀要。325 : 1-218. 土井:10.1206/574.1。hdl : 2246/5999。S2CID 83594819。2012 年 4 月 6 日のオリジナルからアーカイブ (PDF) 。
王暁明。テッドフォード、リチャード H. (2008)。犬: 化石近縁種と進化の歴史。ニューヨーク:コロンビア大学出版局。ISBN 978-0-231-13528-3。OCLC 185095648。
ヤング、SP; ジャクソン、HHT (1978)。賢いコヨーテ。ネブラスカ大学出版局。ISBN 978-0-8032-5893-8。OCLC 3294630。
参考文献編集
本
JS ディクソン (1920 年)。カリフォルニアにおけるコヨーテの防除。カリフォルニア州バークレー : 農業試験場
フローレス、D. (2016)。コヨーテ アメリカ: 自然と超常現象の歴史。基本的な本。
ISBN 978-0-465-05299-8
アーカンソー州ハーディング (1909 年)。オオカミとコヨーテの罠。最新のオオカミ猟師ガイドには、経験豊富な「オオカミ猟師」がこれらの動物を狩り、罠にかける最も成功した方法が記載されており、彼らの習性も詳細に記載されています。オハイオ州コロンバス、アーカンソー州ハーディングのパブ。株式会社
クルテン、B (1974)。「コヨーテに似た犬の歴史(イヌ科、哺乳綱)」。アクタ ズーロジカ フェニカ。140:1-38。
フランソワ・レイデ (1988)。コヨーテ:西部の反抗的なソンドッグ。オクラホマ大学出版局。ISBN 978-0-8061-2123-9。OCLC 17106424。
モーリー、ポール (2004)。シカゴ都市圏における景観利用とコヨーテ Canis latrans の食性(論文)。ユタ州立大学。
ミュリー、A. (1940)。イエローストーンのコヨーテの生態。ワシントン DC : USGPO
パーカー、ジェリー。(1995年)。「Eastern Coyote: Story of Its Success」、Nimbus Publishing、ハリファックス、ノバスコシア州、カナダ。
フランク・ヴァン・ナイズ (2015)。害虫と被害者: アメリカ西部における捕食者規制。カンザス州ローレンス:カンザス大学出版局。
ワーグナー、MM (1920 年頃)。飼いならされたコヨーテの自伝。サンフランシスコ、ハー・ワグナー・パブ。株式会社
ビデオ
シェリー、プリヤ。コヨーテと暮らす(18 分)。イオン。
オーディオブック
ジャック・オルソン。ラスト・コヨーテ(8時間)。ナレーションはゲイリー・マクファーデン。当初は、「Saughter the Animals, Poison the Earth」、Simon & Schuster、1971 年 10 月 11 日に出版されました。ASIN B00WGUA5HK。
外部リンク
![]()
ウィキクォートにコヨーテ
に関する引用句が
![]()
ウィキスピーシーズにはイヌ・ラトランスに関する情報が
![]()
無料の辞書ウィクショナリーで
コヨーテ![]()
・コモンズには、コヨーテに関連するメディアが
「コヨーテ」 。ブリタニカ百科事典。Vol. 7(第11版)。1911年。
「イヌ・ラトランス」。統合分類情報システム。2006 年3 月 23 日に取得。
アリゾナ狩猟魚部門「コヨーテとの暮らし」
ウエスタンコヨーテ 2017 年 9 月 22 日、トレント大学のウルフとコヨーテの DNA バンク、ウェイバック マシンでアーカイブ
生物多様性遺産ライブラリーでイヌ・ラトランスの出現を見る · “