Ribonucleotide
生化学では、リボヌクレオチドは、ペントース成分としてリボースを含むヌクレオチドです。これは核酸の分子前駆体と考えられています。ヌクレオチドは、 DNAとRNAの基本的な構成要素です。リボヌクレオチド自体は、RNA の基本的なモノマー構成要素です。デオキシリボヌクレオチドは、リボヌクレオチド レダクターゼ(RNR)酵素によってリボヌクレオチドを還元することによって形成され、DNA の必須の構成要素です。 DNA デオキシリボヌクレオチドと RNA リボヌクレオチドの間にはいくつかの違いが連続するヌクレオチドは、ホスホジエステル結合を介して結合されます。
リボヌクレオチドは他の細胞機能にも利用されます。これらの特殊なモノマーは、アデノシン一リン酸 ( AMP )に見られるように、細胞制御と細胞シグナル伝達の両方に利用されます。さらに、リボヌクレオチドは、生物内のエネルギー通貨であるアデノシン三リン酸 ( ATP )に変換できます。リボヌクレオチドは環状アデノシン一リン酸 (サイクリック AMP )に変換されて、生物体内でもホルモンを調節します。生物において、リボヌクレオチドの最も一般的な塩基は、アデニン(A)、グアニン(G)、シトシン(C)、またはウラシル(U) です。窒素含有塩基は、プリンとピリミジンの 2 つの親化合物に分類されます。
一般的なリボヌクレオチド構造: リン酸基、リボース、核酸塩基。
コンテンツ
1 構造
1.1 一般的な構造 1.2 DNA デオキシリボヌクレオチドと RNA リボヌクレオチドの比較 1.3 連続するヌクレオチドを連結する
2 関数
2.1 デオキシリボヌクレオチドの前駆体 2.2 リボヌクレオチドの識別
3 合成
3.1 リボヌクレオチド合成
4 リボヌクレオチドのプレバイオティック合成
5 歴史
6 こちらも参照
7 参考文献
構造
一般的な構造
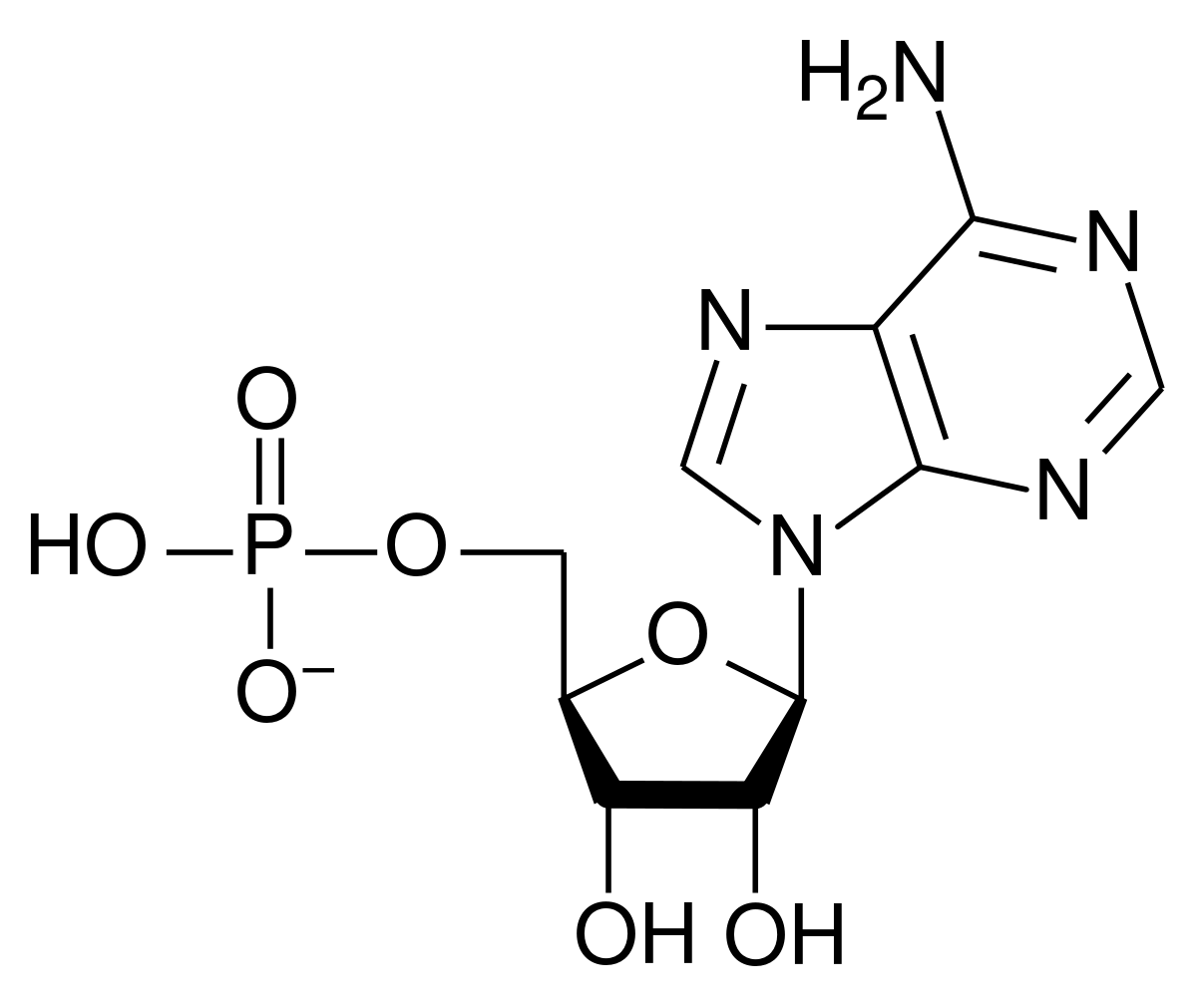
アデノシン 5′-一リン酸 (AMP) の構造

グアノシン 5′-一リン酸 (GMP) の構造

ウリジン 5′-一リン酸 (UMP) の構造

シチジン 5′-一リン酸 (CMP) の構造
リボヌクレオチドの一般的な構造は、リン酸基、リボース糖基、および核酸塩基で構成され、核酸塩基はアデニン、グアニン、シトシン、またはウラシルのいずれかになります。リン酸基のない核酸塩基と糖の組成はヌクレオシドとして知られています。交換可能な窒素含有核酸塩基は、2 つの親化合物、プリンとピリミジンに由来します。ヌクレオチドは複素環式化合物です。つまり、ヌクレオチドは環のメンバーとして少なくとも 2 つの異なる化学元素を含みます。
RNA と DNA の両方には、2 つの主要なプリン塩基、アデニン (A) とグアニン (G) と 2 つの主要なピリミジンが含まれています。DNA と RNA の両方において、ピリミジンの 1 つはシトシン (C) です。ただし、DNA と RNA では 2 番目に重要なピリミジンが異なります。DNA にはチミン (T) が含まれ、RNA にはウラシル (U) が含まれます。まれに、チミンが RNA に存在し、ウラシルが DNA に存在するケースが
ここでは、RNA の構造単位である 4 つの主要なリボヌクレオチド (リボヌクレオシド 5′-一リン酸) を示します。
ヌクレオチド
記号
ヌクレオシド
アデニレート (アデノシン 5′-一リン酸)
あ、アンプ
アデノシン
グアニル酸 (グアノシン 5′-一リン酸) G、GMP グアノシン
ウリジル酸 (ウリジン 5′-一リン酸)
う、UMP
ウリジン
シチジル酸(シチジン 5′-一リン酸) C、CMP シチジン
DNA デオキシリボヌクレオチドと RNA リボヌクレオチドの比較
リボヌクレオチドでは糖成分はリボースですが、デオキシリボヌクレオチドでは糖成分はデオキシリボースです。リボース環の 2 番目の炭素にあるヒドロキシル基の代わりに、水素原子が置換されます。
DNA と RNA のどちらのタイプのペントースも β-フラノース (閉じた 5 員環) の形をしており、核酸の正体を定義します。DNA は 2′-デオキシ-リボース核酸を含むことによって定義され、RNA はリボース核酸を含むことによって定義されます。
場合によっては、DNA および RNA に少量の塩基が含まれる場合がDNA では主要な塩基のメチル化された形態が最も一般的です。ウイルス DNA では、一部の塩基がヒドロキシメチル化またはグルコシル化されている場合がRNA では、マイナー塩基または修飾塩基がより頻繁に発生します。いくつかの例には、ヒポキサンチン、ジヒドロロウラシル、ウラシルのメチル化形態、シトシン、グアニン、および修飾ヌクレオシド プソイドウリジンが含まれます。 5′ 炭素以外の位置にリン酸基を持つヌクレオチドも観察されています。例には、単離可能な中間体であるリボヌクレオシド 2’,3′-環状一リン酸、および特定のリボヌクレアーゼによる RNA の加水分解の最終生成物であるリボヌクレオシド 3′-一リン酸が含まれます。他のバリエーションには、アデノシン 3’,5′-環状一リン酸 (cAMP) およびグアノシン 3’,5′-環状一リン酸 (cGMP) が含まれます。
連続するヌクレオチドを連結する
リボヌクレオチドはホスホジエステル結合を介して結合して RNA 鎖を形成します。1 つのヌクレオチドの 5′-リン酸基は次のヌクレオチドの 3′-ヒドロキシル基に結合し、リン酸残基とペントース残基が交互に並ぶ骨格を形成します。ポリヌクレオチドの各末端にはホスホジエステル結合はありません。酵素RNA ポリメラーゼによってリボヌクレオチド間にホスホジエステル結合が形成されます。RNA 鎖は、鎖の最後のリボヌクレオチドの 3′-ヒドロキシル基が求核剤として作用し、入ってくるリボヌクレオチドの 5′-三リン酸に対して親水性攻撃を開始して、5′ 末端から 3′ 末端まで合成されます。副生成物としてのピロリン酸塩。ヌクレオチドの物理的特性により、RNA の主鎖は非常に親水性が高く極性が中性 pH では、各リン酸基が負の電荷を帯びているため、核酸は高度に帯電します。
DNA と RNA は両方とも、モノヌクレオチド モノマーとしても知られるヌクレオシド リン酸から構築されており、アミノ酸よりも熱力学的に結合する可能性が低くなります。ホスホジエステル結合は加水分解されると、かなりの量の自由エネルギーを放出します。したがって、核酸は自発的に加水分解してモノヌクレオチドになる傾向がRNA の前駆体は GTP、CTP、UTP、および ATP であり、基転移反応における主要なエネルギー源です。
関数
デオキシリボヌクレオチドの前駆体
科学者は、RNA は DNA よりも前に開発されたと考えています。
リボヌクレオチドのデオキシリボヌクレオチドへの還元は、リボヌクレオチド還元酵素によって触媒されます。リボヌクレオチド レダクターゼ (RNR) は、DNA の複製と修復に必要な 4 つのデオキシリボヌクレオチド (dNTP) の合成の最終段階を担うため、すべての生物にとって必須の酵素です。この反応には、他の 2 つのタンパク質、チオレドキシンとチオレドキシン レダクターゼも必要です。リボヌクレオシド二リン酸 (NDP) は、チオレドキシンによってデオキシリボヌクレオシド二リン酸 (dNTP) に還元されます。
一般的な反応は次のとおりです: リボヌクレオシド二リン酸 + NADPH + H + -> デオキシリボヌクレオシド二リン酸 + NADP + + H 2 O
この式を説明するために、dATP と dGTP はそれぞれ ADP と GDP から合成されます。これらはまず RNR によって還元され、次にヌクレオシド二リン酸キナーゼによってリン酸化されて dATP と dGTP になります。リボヌクレオチド還元酵素は、アロステリック相互作用によって制御されます。dATP がリボヌクレオチド レダクターゼに結合すると、デオキシリボヌクレオチドが豊富に存在することを意味するため、酵素の全体的な触媒活性が低下します。このフィードバック阻害は、ATP が結合すると元に戻ります。
リボヌクレオチドの識別
DNA合成中、DNAポリメラーゼは、デオキシリボヌクレオチドと比較してはるかに高いレベルで存在するリボヌクレオチドを選択しなければなりません。生物のゲノムを維持するには DNA 複製が正確である必要があるため、選択性があることが重要です。Y ファミリー DNA ポリメラーゼの活性部位は、リボヌクレオチドに対する高い選択性の維持に関与していることが示されています。ほとんどの DNA ポリメラーゼは、リボース環の 2′-ヒドロキシル基を立体的にブロックできるかさばる側鎖残基を介して、活性部位からリボヌクレオチドを排除する機能も備えています。しかし、多くの核複製および修復 DNA ポリメラーゼはリボヌクレオチドを DNA に取り込むため 、排除機構が完全ではないことが示唆されています。
合成
リボヌクレオチド合成
リボヌクレオチドは、生物体内でより小さな分子からデノボ経路を通じて合成されることも、サルベージ経路を通じてリサイクルされることもある。de novo 経路の場合、プリンとピリミジンは両方とも、アミノ酸の前駆体、リボース-5-リン酸、CO 2、および NH 3に由来する成分から合成されます。

IMPの合成。カラースキームは次のとおりです:酵素、補酵素、基質名、金属イオン、無機分子
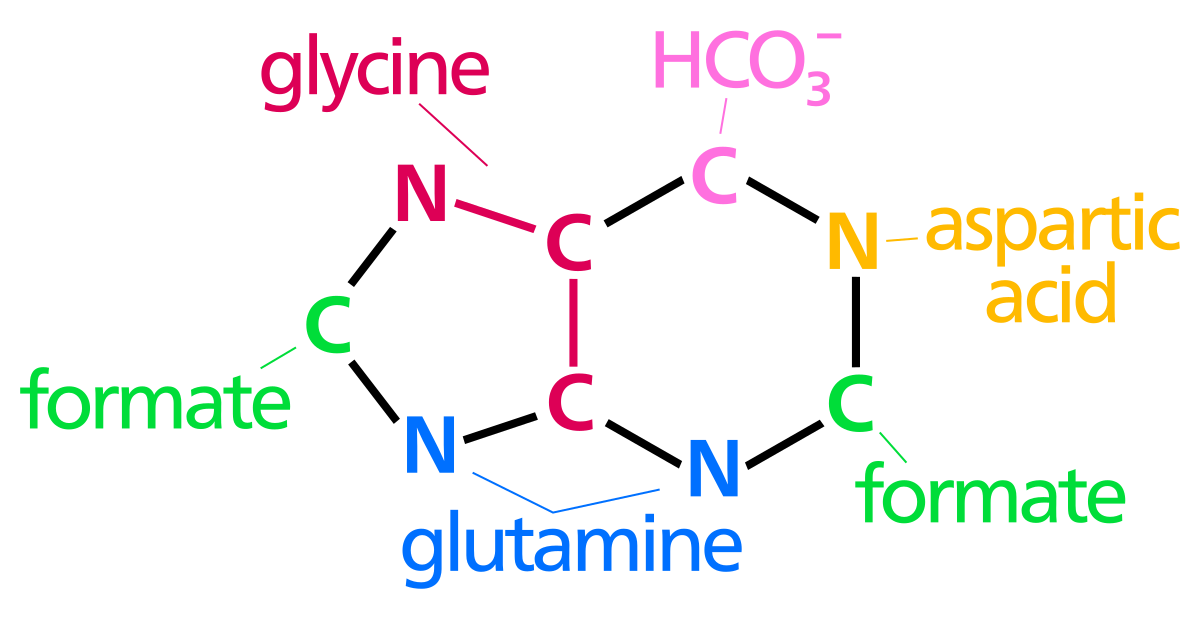
プリン環原子の生合成起源N 1はAspのアミン基に由来しますC 2および C 8 はギ酸に由来しますN 3および N 9はGlnのアミド基によって寄与されますC 4、C 5および N 7はGly Cに由来します6は HCO 3 − (CO 2 )から来ます
プリンヌクレオチドのデノボ生合成はかなり複雑であり、いくつかの酵素反応から構成されます。糖の五環構造を基礎として利用し、プリン環は 11 段階のプロセスで一度に数原子ずつ構築され、イノシン酸 (IMP) の形成につながります。基本的に、IMP は核酸合成に必要なプリンヌクレオチドに変換されます。
この経路は、酵素リボース-リン酸ジホスホキナーゼ (PRPS1) によるリボース-5-リン酸 (R5P) のホスホリボシル ピロリン酸 (PRPP) への変換から始まります。次に、グルタミンが PRPP の C-1 にアミノ基を供与するため、PRPP は 5-ホスホリボシルアミン (5-PRA) に変換されます。縮合反応では、酵素 GAR シンテターゼがグリシンおよび ATP とともに 5-PRA のグリシン カルボキシラーゼ グループを活性化し、グリシンアミドリボヌクレオチド (GAR) を形成します。補酵素 N10-ホルミル-THF は、酵素 GAR トランスフォルミラーゼとともに、GAR のグリシン上のアミノ基に 1 炭素単位を供与し、続いて酵素 FGAR アミドトランスフェラーゼによってグルタミンが付加され、ホルミルグリシンアミジン リボヌクレオチド (FGAM) が形成されます。 )。酵素 FGAM シクラーゼによる FGAM の脱水により、5-アミノイミダゾール リボヌクレオチド (AIR) としてイミダゾール環が閉じられます。N5-CAIR シンテターゼによってカルボキシル基が AIR に結合されて N5-カルボキシアミノイミダゾール リボヌクレオチド (N5-CAIR) が形成され、これが酵素 N5-CAIR ムターゼによってカルボキシアミノイミダゾール リボヌクレオチド (CAIR) に変換されます。酵素 SAICAR シンテターゼは、アスパラギン酸のアミノ基とともにアミド結合を形成し、N-スクシニル-5-アミノイミダゾール-4-カルボキサミド リボヌクレオチド (SAICAR) を生成します。経路をさらに下に進むと、SAICAR リアーゼによるアスパラギン酸の炭素骨格の除去により、5-アミノイミダゾール-4-カルボキサミド リボヌクレオチド (AICAR) が生成されます。酵素 AICAR トランスフォルミラーゼは、N10-ホルミルテトラヒドロ葉酸からの最終的な炭素移動を助け、N-ホルミルアミノイミダゾール-4-カルボキサミド リボヌクレオチド (FAICAR) を形成します。最後に、第 2 の環構造の閉鎖が IMP シンターゼによって行われ、IMP が形成されます。IMP の運命はプリン ヌクレオチドの形成につながります。

ピリミジン de Novo 経路
ピリミジン ヌクレオチドの合成は、はるかに単純なプロセスです。ピリミジン環の形成は、カルバモイルリン酸との縮合反応によるアスパラギン酸のN-カルバモイルアスパラギン酸への変換から始まります。次に、ジヒドロオロターゼおよびジヒドロオロターゼデヒドロゲナーゼは、N-カルバモイルアスパラギン酸をオロチン酸に変換します。オロチン酸は、オロチン酸ホスホリビソールトランスフェラーゼによってホスホリボシルピロリン酸(PRPP)と共有結合し、オロチジン一リン酸(OMP)を生成します。OMP は、オロチジル酸デカルボキシラーゼによる脱炭酸を経て、ウリジル酸 (UMP) リボヌクレオチド構造を形成します。次に、UMP は 2 つのキナーゼ反応によってウリジン-5′-三リン酸 (UTP) に変換されます。UTP からのシチジン-5′-三リン酸 (CTP) の形成は、シチジル酸シンテターゼによるアシルリン酸中間体によって達成できます。
リボヌクレオチドのプレバイオティック合成
生命がどのように誕生したかを理解するには、もっともらしい前生物的条件下で生命の重要な構成要素の形成を可能にする化学経路に関する知識が必要です。RNAワールドの仮説によれば、原始的なスープには浮遊リボヌクレオチドが存在していました。これらは、直列に結合してRNAを形成する基本的な分子でした。RNA と同じくらい複雑な分子は、その反応性が物理化学的プロセスによって支配される小分子から生じたに違いありません。RNAはプリンとピリミジンのヌクレオチドで構成されており、どちらも信頼性の高い情報伝達、ひいてはダーウィンの自然選択と進化に必要です。活性化ピリミジンリボヌクレオチドの合成は、もっともらしいプレバイオティック条件下で実証されました。合成の出発物質 (シアナミド、シアノアセチレン、グリコールアルデヒド、グリセルアルデヒド、無機リン酸塩) は、プレバイオティクス原料分子として考えられると考えられました。ナムら。 は、核酸塩基とリボースが直接縮合して、水性微液滴中でリボヌクレオシドを生成することを実証しました。これは、RNA 形成につながる重要なステップです。また、湿式-乾式サイクルを使用してピリミジンおよびプリンのリボヌクレオチドを合成するためのもっともらしいプレバイオティクスプロセスが、Becker et al. によって提示されました。
歴史
フィーバス・レヴィン
ロザリンド・フランクリンのX線結晶構造解析画像からDNAの構造を詳細に説明したジェームズ・ワトソンとフランシス・クリックの画期的な論文以前にも、その発見に貢献した歴史的な科学者が何人かいた。フリードリヒ・ミーッシャーはスイスの医師で、1869年に白血球の核から初めて核物質を単離・同定し、後に「ヌクレイン」と呼び、DNA発見への道を開いた。ミーッシャーの研究に続いて、ドイツの生化学者アルブレヒト・コッセルが 1878 年に「ヌクレイン」の非タンパク質成分を単離し、核酸に存在する 5 つの核酸塩基、アデニン、シトシン、グアニン、チミン、ウラシルを発見しました。 。これらの初期の発見により、核酸に関するいくつかの基本的な事実が知られていましたが、その構造と機能は謎のままでした。
1919 年にロシア系リトアニア人の生化学者フィーバス・レベンによってヌクレオチドが発見されて初めて、DNA 発見の扉が再び開かれました。レベンは、酵母のRNAに存在する炭水化物成分が実際にはリボースであることを最初に特定しました。しかし、胸腺核酸の炭水化物成分も糖であるが、デオキシリボースと呼ばれる酸素原子が 1 つ欠如していることを発見して初めて、彼の発見は科学界に広く評価されました。最終的に、レベンは、RNA と DNA の構成要素が組み合わされる正しい順序、つまりリン酸塩 – 糖塩基 – 塩基単位を特定することができました。これを彼は後にヌクレオチドと呼びました。レヴィンはヌクレオチド成分の順序をよく理解していましたが、宇宙におけるヌクレオチド配列の構造とその遺伝暗号は、彼のキャリアの初期には依然として謎のままでした。
こちらも参照
リボヌクレオシドまたはリボシド
参考文献
^ ネルソン、デイヴィッド (2008). レーニンガー生化学の原理。WH Freeman and Co.、272 ~ 273 ページ。
^ ニュースホルム、エリック A.; リーチ、アンソニー R. ボード、メアリー (2008)。健康と病気における機能生化学: 健康と病気における代謝制御(第 2 版)。ニュージャージー州ホーボーケン: ワイリー。ISBN 978-0-471-98820-5。
^ ダス、デバジョティ (2010). 生化学。Academic Publishers の Bimal Kumar Dhur 氏。
^ コックス、マイケル M. ネルソン、デイビッド L. (2008)。生化学の原理。WH フリーマン&カンパニーISBN 978-1-4292-2263-1。
^ レイモンド、ケネス W. (2010)。一般化学、有機化学、生物化学: 統合的アプローチ(第 3 版)。ニュージャージー州ホーボーケン: ワイリー。ISBN 978-0-470-55124-0。
^ シェヒター、モセリオ; ジョシュア・レーダーバーグ編。(2004)。卓上微生物百科事典(第 1 版)。アムステルダム: Elsevier Acad. プレス。ISBN 0-12-621361-5。
^ フィル、ターナー; 他。(2005)。分子生物学。インスタント ノート (第 3 版)。フロリダ州ボカラトン: CRC、テイラー & フランシス。ISBN 0-415-35167-7。
^ ネルソン、デヴィッド (2008). レーニンガー生化学の原理。WH Freeman and Co.、274 ~ 275 ページ。
^ チョーハン、アショク K.; ヴァルマ、アジット編。(2009年)。分子バイオテクノロジーの教科書。ニューデリー: IK International Pub. 家。ISBN 978-93-80026-37-4。
^ チェンドラ・ムデル、M; フアレス、A; トレント、E (2012)。「バイオフィルムは大腸菌におけるリボヌクレオチド還元酵素遺伝子の発現を変化させる」。プロスワン。7 (9): e46350。Bibcode : 2012PLoSO…746350C。土井:10.1371/journal.pone.0046350。PMC 3458845。PMID 23050019。 ^ メアリー K. キャンベル; ファレル、ショーン O. (2009)。生化学(第 7 版)。カリフォルニア州ベルモント: ブルックス/コール・センゲージ学習。ISBN 978-0-8400-6858-3。
^ バーグ、ジェレミー M. ティモチコ、ジョン L. ルバート・ストリヤー (2007)。生化学(第 6 版、第 3 刷版)。ニューヨーク:フリーマン。ISBN 978-0-7167-8724-2。
^ ケビン・N・キルアック、ズカイ・スオ、ホン・リン、ケビン・N. 周防、図彩。リン、ホン(2011 年 4 月 1 日)「YファミリーDNAポリメラーゼによるリボヌクレオチド識別の構造機構」。分子生物学ジャーナル。407 (3): 382–390。土井:10.1016/j.jmb.2011.01.037。PMID 21295588。 ^ ニック・マケルヒニー、SA; クマール、D; アラバマ州クラーク。ワット、DL; ワシントン州ワッツ。ルンドストローム、EB; ヨハンソン、E; チャベス、A; TA州クンケル。「DNAへのリボヌクレオチドの取り込みによるゲノムの不安定性」。自然のケミカルバイオロジー。6 (10): 774–81。土井:10.1038/nchembio.424。PMC 2942972。PMID 20729855。 ^ ニック・マケルヒニー、SA; ワシントン州ワッツ。クマール、D; ワット、DL; ルンドストローム、EB; バーガーズ首相。ヨハンソン、E; チャベス、A; TA州クンケル(2010年3月16日)。「酵母の複製ポリメラーゼによるDNAへの豊富なリボヌクレオチドの取り込み」。アメリカ合衆国国立科学アカデミーの議事録。107 (11): 4949–54。Bibcode : 2010PNAS..107.4949N。土井:10.1073/pnas.0914857107。PMC 2841928。PMID 20194773。 ^ カシヴィスワナタン、R; トイレットペーパーのコープランド(2011 年 9 月 9 日)。「ヒトミトコンドリアDNAポリメラーゼによるリボヌクレオチドの識別と逆転写」。生物化学ジャーナル。286 (36): 31490–500。土井: 10.1074/jbc.M111.252460。PMC 3173122。PMID 21778232。 ^ ネルソン、デイヴィッド (2008). レーニンガー生化学の原理。WH Freeman and Co.、881 ~ 894 ページ。
^ バーグ、JM (2002). 生化学。プリン塩基は、de Novo によって合成することも、Salvage Pathways によってリサイクルすることもできます。ニューヨーク:WHフリーマン。pp. セクション 25.2。
^ オーナー MW、ガーランド B、サザーランド JD。プレバイオティクス的に妥当な条件での活性化ピリミジンリボヌクレオチドの合成。自然。2009 年 5 月 14 日;459(7244):239-42。土井:10.1038/nature08013。PMID: 19444213 ^ ナムI、ナムHG、ザレRN。水性微液滴中でのプリンおよびピリミジンリボヌクレオシドの非生物的合成。Proc Natl Acad Sci US A. 2018 1 2;115(1):36-40。土井: 10.1073/pnas.1718559115。Epub 2017 12 18. PMID: 29255025; PMCID: PMC5776833 ^ Becker S、Feldmann J、Wiedemann S、Okamura H、Schneider C、Iwan K、Crisp A、Rossa M、Amatov T、Carell T. プレバイオティクスとして妥当なピリミジンおよびプリン RNA リボヌクレオチドの統合合成。化学。2019 10 4;366(6461):76-82。土井: 10.1126/science.aax2747。PMID: 31604305。
^ ワトソン、JD; クリック、FH (1953 年 4 月 25 日)。「核酸の分子構造;デオキシリボース核酸の構造」。自然。171 (4356): 737–8。Bibcode : 1953Natur.171..737W。土井:10.1038/171737a0。PMID 13054692。S2CID 4253007。 ^ ダーム、R 。「DNAの発見:フリードリヒ・ミーッシャーと核酸研究の初期」。人間の遺伝学。122 (6): 565–81。土井:10.1007/s00439-007-0433-0。PMID 17901982。S2CID 915930。 ^ ジョーンズ、私 (1953 年 9月)。「アルブレヒト・コッセル、伝記スケッチ」。イェール生物学医学ジャーナル。26 (1): 80-97。PMC 2599350。PMID 13103145。 ^ レヴィン、フィーバス (1919)。酵母の核酸の構造。生物化学ジャーナル 40(2)。415–24ページ。 · “