Social_grooming
社会的身だしなみは、人間を含む社会的動物が互いの体や外見をきれいにしたり維持したりする行動です。関連する用語であるアログルーミングは、同じ種のメンバー間の社会的なグルーミングを示します。グルーミングは、大きな社会活動、および近接に住む動物がことができる手段で結合し、社会構造、家族へのリンク、およびビルドcompanionshipsを強化します。社会的身だしなみは、紛争解決、母親の行動、および一部の種の和解の手段としても使用されます。 相互グルーミング 通常、2人の個人間の身だしなみの行為を説明します。多くの場合、社会的な身だしなみ、つがい、または交尾前の活動の一部として行われます。
大人の
アヌビスヒヒが幼体を手入れする
メディアを再生する
オスの猫がメスの子猫を手入れします。
コンテンツ
1 進化の利点
1.1 健康上の利点 1.2 社会構造の強化と関係の構築
1.2.1 ソーシャルボンドの作成と維持
1.2.2 ヒエラルキーと社会構造の強化
1.2.3 好意の交換
1.3 直接的な適応度の結果
2 利他主義
2.1 ハミルトンの法則 2.2 商品として使用
3 社会的グルーミングの個体発生
3.1 グルーミングの一般的な学習と往復 3.2 学習における性別の違い
4 ツールの使用法
5 相互グルーミング
5.1 昆虫 5.2 コウモリ 5.3 霊長類 5.4 鳥 5.5 馬 5.6 牛
6 内分泌作用
6.1 オキシトシン 6.2 ベータエンドルフィン 6.3 糖質コルチコイド受容体 6.4 オピオイド
7 引用された研究に対する批判
8 も参照してください
9 参考文献
10 参考文献
11 外部リンク
進化の利点
社会的な身だしなみ行動がフィットネスを向上させると仮定されているさまざまな提案されたメカニズムがこれらの進化的利点は、病気の伝染の減少とストレスレベルの減少、社会構造の維持、生存の尺度としてのフィットネスの直接的な改善などの健康上の利点の形でもたらされる可能性が
健康上の利点
社会的グルーミングの最も重要なことは生物の健康と衛生を高めることであるのか、それとも社会的グルーミングの社会的側面が同等以上の重要な役割を果たすのかについてしばしば議論されます。伝統的に、社会的グルーミングの主な機能は動物の衛生状態の維持であると考えられています。この声明を裏付ける証拠には、グルーミングは自動グルーミングではアクセスできない身体部分に集中し、身体部分がより重要な社会的またはコミュニケーション機能を持っていたとしても、アログルーミング領域に費やされる時間は大幅に変化しなかったという事実が含まれます。
社会的な身だしなみ行動は、さまざまな種で一連の健康上の利点を引き出すことが示されています。たとえば、グループメンバーのつながりは、ストレッサーの潜在的に有害な影響を軽減する可能性がでマカク、社会的グルーミングは減らすことが証明された心拍数を。軽度のストレッサー中の社会的所属は、ラットの乳腺腫瘍発生レベルの低下および寿命の延長と相関することが示されましたが、この所属の欠如が主要な危険因子であることが示されました。グルーミングは、野生のヒヒ(Papio cynocephalus)のダニの負荷を減らすのに不可欠な役割を果たすことも示されています。これらの外部寄生ダニはとして作用する可能性運ぶベクトルの拡散のための疾患及び感染症の一般的なダニ媒介によって寄生虫のようなhaemoprotozoanを。ダニの負荷が低いヒヒは、そのような感染の発生が減少し、ヘマトクリット値(濃厚赤血球の量)が高いことから明らかなように、健康状態が高い兆候を示します。

ダニのために
グルーミングするマントヒヒ
一方で、グルーミングの衛生的側面は、社会的側面ほど重要な役割を果たしていないという議論もあり得ます。44の異なる霊長類の種で行われた観察研究は、種のアログルームが平均して、その体のサイズではなく、そのグループのサイズと相関していることを示唆しています。衛生的な観点から純粋にグルーミングが必要な場合、動物が大きくなればなるほど、そのグループのメンバーによってグルーミングされることが多くなります。ただし、代わりに、グループのサイズが大きくなると、メンバーは全員のグルーミングに適切な時間を費やすようになります。したがって、動物、特にここの霊長類が、衛生的な観点から必要以上に頻繁にお互いをグルーミングするという事実は、アログルーミングの社会的角度が、それ以上ではないにしても、等しく重要な役割を果たしていることを示唆しています。社会的側面の重要性を示すもう1つの証拠は、霊長類が自分自身をグルーミングする量と方法(自動グルーミング)と比較して、アログルーミングにはより長い期間とさまざまな技術が含まれ、その一部には愛情のこもったジェスチャーであるという意味が
社会構造の強化と関係の構築
ソーシャルボンドの作成と維持
ソーシャルグルーミングの最も重要な機能の1つは、ソーシャルネットワークと関係を確立することです。多くの種では、個人は「友情」と呼ばれる密接な社会的つながりを形成します。これは、一緒に活動を行うのに長い時間がかかるためです。特に霊長類では、身だしなみはこれらの友情の形成と維持において大きな社会的重要性と機能を持っていることが知られています。研究は上で実行アカゲザルことを示したのfMRIサルの脳のスキャンがでより顕著に点灯perirhinial皮質及び(認識および記憶に関連する)時間極サルであった場合(社会的および感情的な処理/解析に関連します)あまり馴染みのない顔と比較して、友達の顔の写真を表示しました。したがって、霊長類は、親しみやすく好意的な個人(「友人」)を認識し、あまり好まれていないパートナーと比較して、彼らの身だしなみに多くの時間を費やします。バーバリーマカクのように、より寛容な社会的スタイルを持つ種では、女性は社会的ランクではなく、誰をよく知っているかに基づいてグルーミングメイトを選択することが見られます。霊長類に加えて、鹿、牛、馬、ハタネズミ、ネズミ、ミーアカット、ハナグマ、ライオン、鳥、コウモリなどの動物も、手入れをすることで社会的絆を形成します。

緊密なクラスターのインドの短い鼻のフルーツコウモリ
社会的な身だしなみはまた、仲間や好色なパートナーを確立し、認識するのに役立つかもしれません。たとえば、鼻の短いコウモリでは、雌は夕暮れの飛行の直前に雄と一緒にグルーミングを開始します。オスと彼の緊密なメスのハーレムは、お互いに体の分泌物を適用します。これにより、メスの生殖状態を認識できる可能性が Kumarらによる2016年の研究。これらの分泌物を化学的に分析し、化学感覚を介したコミュニケーションおよび配偶者選択に必要である可能性があることを明らかにしました。同様に、攻撃性の低いハーブフィールドマウス種では、オスはメスをより長い期間グルーミングし、メスが往復しないようにすることさえ観察されます。オスの交配要求はメスが提供するものよりも大きいため、メスは交配を選択する方法としてソーシャルグルーミングを使用し、オスはそれを使用して交配を促します。
最後に、血縁淘汰は、以前考えられていたように、グルーミングメイトを選ぶときの友情やメイトの好みほど重要な要素ではありません。 2018年にフェルプスら。チンパンジーに関する飼育下の研究では、動物は「成功した」または「失敗した」相互作用を覚えており、それをグルーミング仲間を選択するための基礎として使用したことがわかりました。彼らは、誰が往復するかではなく、誰が往復するかに基づいてグルーミング仲間を選びました。さらに重要なことに、2匹のチンパンジーが互いにグルーミングする間の遅延が非常に小さい場合、チンパンジーは「タイムマッチ」する傾向がつまり、2番目のチンパンジーが最初のチンパンジーをグルーミングした時間と同じ時間グルーミングします。この「エピソード記憶」は、要求の厳しい量の認知機能と感情認識を必要とし、食物の好みに関して実験的にテストされています。食べ物を試した後。したがって、類人猿は、さまざまな時間に発生したさまざまなイベントを区別できます。
ヒエラルキーと社会構造の強化
一般に、ソーシャルグルーミングは、階層を上に向けたアクティビティです。つまり、ランクの低い個人は、グループ内のランクの高い個人をグルーミングします。ではミーアキャット、社会的グルーミングは増加の関係維持の役割を運ぶことが示されているフィットネス。このシステムでは、研究者は、優勢な男性はより多くのグルーミングを受け、他の男性はより少なくグルーミングすることを観察しました。アカゲザルを対象に実施された研究では、下位のグループメンバーと比較して、より優勢なグループメンバーが、手入れをするときに「摘み取られた」よりも「撫でられた」ことがわかりました。功利主義の観点からは、撫でることは身だしなみを整えるよりも効果的なテクニックではありませんが、より愛情のこもったジェスチャーであると解釈されます。したがって、潜在的な攻撃者をなだめ、緊張を和らげるために、上位の個人をグルーミングすることができます。さらに、ランクが近い個人は、ランクが遠い個人よりも相互に手入れをする傾向が
カンムリテナガザルのグルーミングネットワークは、より大きな社会的結束と安定に貢献することが証明されています。より安定したソーシャルネットワークを持つテナガザルのグループは、非常に複雑なグルーミングネットワークを形成しましたが、安定性の低いネットワークを持つグループは、はるかに少ないグルーミングペアを形成しました。
好意の交換
身だしなみは、特定の行動反応や行動と引き換えに個人によって提供されることがよく社会的なグルーミングは、特に吸血コウモリにとって重要です。なぜなら、彼らは、彼らの食物逆流共有行動を維持するために、食物共有関係を維持する必要があるからです。で中国チベットマカクは、幼児は便宜のために交換することができる貴重な商品として見られています。母親は、母親以外の人が手入れをすることと引き換えに、乳児を短期間扱うことを許可します。中国チベットマカクは、グループ内の乳児の相対的な比率に注目することにより、乳児の価値を測定および認識します。乳児の数が増えると、その「価値」は低下し、母親以外の母親が母親のために行う乳児の取り扱いと引き換えに身だしなみを整える量は減少します。
男性のボノボスでは、身近な人の身だしなみは時間差が大きく(つまり、各人が他の人を身だしなみする期間が等しくない)、相互関係が低下する(つまり、他の人を身だしなみにする可能性がある)ため、身だしなみを交換して感情的な要素を取り入れることをお勧めします予測不可能です)。したがって、個人間にある種の社会的絆が存在することは、より大きな「寛大さ」と個人間の寛容をもたらす。
直接的な適応度の結果
社会的な身だしなみの関係は、さまざまな種に直接的なフィットネスの利点を提供することが証明されています。特に、キイロヒヒ(Papio cynocephalus)の身だしなみは広く研究されており、社会的な身だしなみ行動によって形成された社会的絆の結果として適応度が向上することを示す多くの研究が野生のヒヒに関する16年間の行動データを収集したそのような研究の1つは、社会性が乳児の生存に及ぼす影響を浮き彫りにしています。 1年までの乳児の生存率と、近接性および社会的身だしなみに基づく社会性の尺度である複合社会性指数との間に正の関係が確立されている。野生のヒヒの成虫の生存に対する社会性の影響についての証拠も提供されています。社会的つながりの測定値(社会的身だしなみに焦点を当てる)と雌と雄の両方のヒヒの生存期間の中央値との間の直接的な相関関係がモデル化されました。
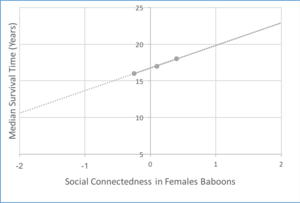
雌ヒヒの生存期間中央値と社会的つながりの関係
身だしなみによって確立された社会的絆は、紛争解決と攻撃からの保護という形で適応的な利点を提供する可能性が野生のサバンナのヒヒでは、社会的所属は、より支配的なグループメンバーからの寛容を高め、グループ内のコンテストの相互作用のインスタンス中に同種から援助を得る機会を増やすことによって、フィットネスを強化することが示されています。キイロヒヒでは、大人の女性が親族と関係を築き、社会集団内での激しい紛争の際に支援を提供します。でバーバリーのマカク、パートナー間の重要な関係の形成における社会的グルーミング結果。これらの社会的関係は、協力を支援し、しばしば身体的危害を引き起こす可能性のある他の男性で構成される戦闘グループに対する保護を促進するのに役立ちます。さらに、社会的関係は、いくつかの霊長類における子殺しのリスクを減少させることも証明されている。
利他主義
利他主義は、生物学的な意味で、行動を実行する人の適応度を低下させながら、別の個人の適応度を増加させる個人によって実行される行動を指します。これは、他者を助けるという意識的な意図を必要とする利他主義の哲学的概念とは異なります。行動として、利他主義は道徳的な観点から評価されるのではなく、生殖適応度のための行動の結果として評価されます。利他主義の明らかな利点の背後にあるメカニズムとして、チャールズ・ダーウィンは群選択を提案しました。
社会的身だしなみは、通性利他主義の行動と見なされます。行動自体は、直接的な適応度の一時的な喪失(間接的な適応度の増加の可能性を伴う)と、それに続く個人的な生殖です。このトレードオフは、囚人のジレンマモデルと比較されており、この比較から、「しっぺ返し」というタイトルのロバートトリヴァース互恵的利他主義理論が生まれました。利他主義と併せて、血縁淘汰は、生物自身の生存と繁殖を犠牲にしても、生物の親戚の繁殖成功を支持することに重点を置いています。このため、血縁淘汰は、生産された子孫の数と、兄弟などの他の人を支援することによって個人が確実に生産できる数を組み合わせた包括的適応度の例です。
ハミルトンの法則編集 >> {rB> C}
C}””>
WDハミルトンによって開発されたこのルールは、俳優に対するレシピエントの遺伝的関連性(r)にレシピエントへの利益(B)を掛けたものが、俳優(C)。したがって、行動の恩恵を受ける個人が行動を提供する個人に関連している限り、個人が社会的グルーミングなどの利他的な行動に参加することは有利である。
商品として使用
一部の動物が代わりに利他的な行動を市場戦略として使用して、望ましいものと交換しているかどうかが疑問視されました。アヌビスヒヒ、Papio anubisでは、攻撃性の低下などの利益と引き換えに行動が提供される取引の一形態として、個人が利他的な行動をとることがわかっています。身だしなみは、単一の発作ではなく複数の発作にわたって均等にバランスが取れていた。これは、女性が単一の取引との完全な交換に制約されず、社会的グルーミングを使用して社会集団の人々との長期的な関係を固めることを示唆している。
さらに、シロテテナガザル(Hylobates lar)は、グループ内のメスの発情時にオスが社会的グルーミングにもっと注意を払っていることを確認しました。社会的グルーミング自体の行動は、サービスを提供する人にとって有益ではありませんでしたが、交尾とその後の受精の機会は、行動に参加する人々の生殖適応度を高めます。この研究はまた、社会的グルーミングのパフォーマンスが女性の卵巣サイクルのパフォーマンスと循環していることを発見することに成功しました。
社会的グルーミングの個体発生
グルーミングの一般的な学習と往復
ほとんどの場合、グルーミングは個人の母親から学ぶ行動です。乳児は母親によって手入れされており、これらの行動をお互いに、そして母親を幼い頃に模倣している。この行動は、個人が完全に発達した成人になり、通常のグルーミングパターンに従うことができるようになると、他のグループメンバー(母親以外または異なるランク)でより頻繁に往復します。
学習における性別の違い
ある種のオスとメスのメンバーは、どのように、いつ、誰を手入れするかを学ぶ点で異なる場合がで切り株尾マカク、乳児の女性がカウンターパートが行う彼らの男性よりも頻繁に母親をグルーミングすることにより、その母親が新郎と同じグループのメンバーをグルーミングによって母親の行動を模倣します。この擬態は、乳児のベニガオザルにおける識別に基づく観察学習を示すことが示唆されており、母親の擬態および親族に偏ったグルーミングに対する娘のペンチと、ランクに偏ったグルーミングに対する息子のペンチは、彼らの社会と一致している成人男性がランクを獲得して維持するために同盟を必要とするグループでの役割。
ツールの使用法
ソーシャルグルーミングのほぼすべての例で、個人は手、歯、舌などの自分の体の部分を使用して、グループのメンバーや幼児をグルーミングします。人間以外の動物の社会的グルーミングで道具の使用例を観察することは非常にまれです。しかし、霊長類ではそのような例がいくつか観察されています。1981年にバックネル大学で行われたニホンザルの観察研究では、母親のマカクが地面にあるいくつかの石を観察した後、石を選び、この石を使って乳児を手入れしているのが見られました。石は乳児の気を散らすものとして使用され、母親が他の場所で注意を向けている間に乳児を適切にきれいにすることができると仮定された。これは、母親が石を落とした後、乳児が石を拾い上げ、遊んでいる間、彼女に手入れをさせたという事実によって裏付けられました。この行動は、コロニー内の他の数人のメンバーで見られましたが、種全体ではまったく見られませんでした。別の例では、デルタ地域霊長類研究センターのメスのチンパンジーが、葉の小枝を剥がして「歯ブラシ」を作成し、この歯ブラシを使用して、いくつかの例で乳児を手入れしました。しかしながら、両方の例は霊長目での道具の使用に関係しており、それはすでに広く研究され、科学的に裏付けられています。霊長類の広い作業記憶容量と因果関係の理解能力により、霊長類は他の非人間動物よりもはるかに広範囲に道具を作り、利用することができます。肉体的および精神的制約は別として、おそらく動物のグルーミングが道具を使用しない理由は、社会的グルーミングの主な目的が社会的結合であり、感情的な交換を伴うためであり、その多くは触覚によって伝えられます。
相互グルーミング

ポニーの相互グルーミング

お互いをグルーミングする3匹のマカク

スミレコンゴウインコの社会的グルーミング
 メディアを再生する
メディアを再生する
女性セキセイインコのオス(ビデオ)を修復します
 メディアを再生する
メディアを再生する
あるライオンが別のライオンをグルーミングする

でAllopreening黄色いくちばしおしゃべり

ボンネットマカクが乳児の乳児を吸っている間、身だしなみを整える
多くの動物は、撫でたり、引っ掻いたり、マッサージしたりする形でお互いを手入れします。この活動は、これらの社会的に活動的な動物の共同の成功を促進するために、体から異物を取り除くのに役立つことがよく霊長類、昆虫、鳥、、コウモリなど、王国全体にさまざまな社会的グルーミング動物が存在します。徹底的な研究はまだ行われていないが、霊長類の研究を通じて、人間以外の動物の社会的グルーミングについて多くのことが学ばれている。哺乳類の社会的身だしなみの背後にある原動力は、主に、食物、性別、共同衛生などの資源の交換における功利主義的な目的だけでなく、慰めの行動への適応に根ざしていると考えられています。

昆虫
昆虫では、手入れはしばしば体から異物を取り除くという重要な役割を伴います。ミツバチは、例えば、受信蜂によって到達することができない体の部分を洗浄することにより、社会的グルーミングを行っています。受け入れ蜂は、ほこりや花粉を取り除くために、羽、口の部分、触角を掃除しながら、羽を体に垂直に伸ばします。このほこりや花粉の除去は、グループの全体的な幸福に貢献する嗅覚の研ぎ澄ましを可能にします。
コウモリ

最近の研究によると、吸血コウモリは、グループの幸福を促進するために、他の種類のコウモリよりもはるかに多くの社会的グルーミングに従事していることがわかっています。より高いレベルの寄生虫感染に直面して、吸血コウモリは、逆流を介して食物を共有するだけでなく、お互いを掃除することに従事します。この活動は、グループの成功を促進しながら、進行中の感染を防ぎます。
霊長類

メディアを再生する
ニホンザル(ニホンザルで互いにグルーミング)
地獄谷モンキーパークで
山之内
霊長類は、彼らのさまざまなライフスタイルに関して行われた集中的な研究、および異なる種間での社会的グルーミングの手段の直接的な変化のために、おそらく相互グルーミングの最良の例の1つを提供します。霊長類の間では、社会的グルーミングは動物の慰め行動において重要な役割を果たし、霊長類は支配階層、既存の連合を通じて、そして紛争後の和解のために同盟を確立し維持することに従事します。霊長類は退屈の瞬間にも社会的に手入れをし、その行為は緊張とストレスを軽減することが示されています。このストレスの減少は、観察されたリラックスした行動の期間と関連していることが多く、霊長類は身づくろいを受けている間に眠りに落ちることが知られています。霊長類間の対立は、グループ間のストレスの増加として研究者によって観察されており、相互グルーミングが非常に有利になっています。
グルーミングを開始することには利点がグルーミングを開始した人は、代わりに自分でグルーミングされ、掃除されるという利点が研究によると、社会的はしごの低い霊長類は、彼らの地位を高めるために、より高いランクの霊長類で身づくろいを開始する可能性が紛争や競争が激化する時期には、これが発生する可能性は低いことがわかっています。霊長類は、身だしなみの使用のバランスをとる必要があると考えているかもしれないと研究者は示唆しています。
霊長類の身づくろいは、同盟の形成と維持のためだけでなく、共同の食物、性別、衛生などの資源を交換するためにも利用されます。野生のヒヒは、ダニや他の昆虫を他の人から取り除く活動として、社会的な身づくろいを利用することがわかっています。このグルーミングでは、注目を集めている体の部分は、ヒヒ自体が届かない部分のようです。これらの地域でのグルーミング活動は、寄生虫、汚れ、死んだ皮膚、およびもつれた毛皮を取り除くために使用され、特定の領域に到達してきれいにすることができないにもかかわらず、動物の健康を良好な状態に保つのに役立ちます。
霊長類がグルーミングに費やす時間はグループのサイズとともに増加しますが、グルーミングに費やす時間は通常他の要因の影響を受けるため、グループのサイズが大きすぎるとグループの凝集性が低下する可能性がその結果、グルーミングに費やされる時間に影響を与えるこれらの側面のいくつかには、生態学的、系統発生的、および生活史が含まれます。たとえば、、「認知的制約と捕食圧力はグループのサイズに強く影響し、それによって霊長類のグルーミング時間に間接的な影響を及ぼします」と述べています。過去のデータとこのトピックに関して行われた研究を分析することにより、著者は40を超える霊長類グループがより大きな生態学的問題に直面し、したがって社会的グルーミングに費やされる時間が影響を受けることを発見した。
チンパンジーに関する最近の研究では、オキシトシンの放出と慰めの行動との直接的な相関関係が明らかになっています。この行動と放出は、幼児期から成体期まで社会的グルーミングに積極的に従事する霊長類種であるベルベットモンキーなどの霊長類で注目されています。ベルベットモンキーの兄弟は、母親による身だしなみの割り当てをめぐって対立することがよくありますが、身だしなみは緊張を仲介する活動であり、同盟の形成と維持に低コストです。この手入れは、兄弟の間だけでなく、母親を巻き込むことの両方で行われます。
最近の研究カニクイザルは、男性が調達セックスするためにメスをグルーミングすることを示しています。ある研究によると、女性は、最近手入れをした場合、手入れをしなかった男性と比較して、男性と性的活動を行う可能性が高いことがわかりました。
鳥
手入れ(鳥)§アロプリーニング
鳥はアロプリーニングに従事します。研究者は、この慣行がつがいを構築すると信じています。 2010年、研究者は、傍観者との接触を介したレイヴン内の慰め行動としての社会的グルーミングの形態の存在を決定しました。くちばしに触れる。
馬
馬は、体の表面の寄生虫やその他の汚染物質が積極的に除去される「つがい」の形成を介して相互グルーミングに従事します。この異物の除去は、主に首などの手の届きにくい場所でニブリングを介して行われます。
牛
グルーミングは、乳製品や牛肉の品種を含む多くの種類の牛に一般的に見られる行動です。社会的なめる行為は、特に未経産牛に見られ、社会的支配を開始し、交際を強調し、自分自身または他者の衛生状態を改善します。牛に見られるこの行動は、寄生虫の負荷の減少、社会的緊張、飼料貯蔵庫での競争などの利点をもたらす可能性が社会的な舐めは、前向きな感情やリラックスした環境を促進するなどの長期的な利益をもたらすことができると理解されています。
内分泌作用
社会的身だしなみは、個人内の内分泌レベルの変化と相関していることが示されています。具体的には、脳からのオキシトシンの放出と社会的身づくろいの間には大きな相関関係がオキシトシンは、放出されたときの肯定的な感情的反応により、向社会的行動を促進すると仮定されています。さらに、社会的グルーミングは、ストレス軽減における生理学的反応を促進するベータエンドルフィンも放出します。これらの反応は、ホルモンやエンドルフィンの産生から、または神経構造の成長や減少を通じて発生する可能性がたとえば、授乳中のラットの研究では、授乳時に温かさと接触を受けたラットは、接触を受けなかったラットよりも血圧レベルが低かった。これは、迷走神経緊張の増加の結果であることがわかりました。つまり、刺激に対する副交感神経反応が高く、交感神経反応が低く、ストレス反応が低くなっています。社会的身だしなみは、無害な感覚の活性化の一形態です。非攻撃的な接触を特徴とする無害な感覚の活性化は、無害な攻撃的な感覚の活性化から完全に分離した神経経路を刺激します。無害な感覚は、後柱-内側の辺縁系を介して伝達されます。
オキシトシン
オキシトシンは、利他主義などの社会的感情を表現するのに役立つことが知られているペプチドホルモンであり、社会的行動に正のフィードバックメカニズムを提供します。例えば、吸血コウモリの研究は、オキシトシンの鼻腔内注射が雌のコウモリによって行われるグルーミングの量を増加させたことを示しました。ポジティブタッチ(グルーミングなど)、ポジティブな匂いや音によって刺激されることがわかっているオキシトシンの放出は、個人に生理学的利益をもたらす可能性が利点には、リラクゼーション、癒し、消化刺激などがさらに、ラットでの研究がオキシトシンの放出が男性の生殖の成功を増加させることができることを示したように、生殖の利益が発見されました。オキシトシンの役割は、母体のつがいにおいて重要であり、社会的相互作用からの正のフィードバックループの結果として、社会的グループにおける同様の結合を促進すると仮定されています。
ベータエンドルフィン
グルーミングはベータエンドルフィンの放出を刺激します。これは、グルーミングがリラックスしているように見える理由の1つの生理学的理由です。ベータエンドルフィンは、視床下部および下垂体のニューロンに見られます。ベータエンドルフィンはオピオイドアゴニストであることがわかっています。オピオイドは、受容体に作用してリラックス感を促進し、痛みを軽減する分子です。サルでの研究は、ベータエンドルフィンレベルの変化を反映した、体内のアヘン剤発現の変化が社会的グルーミングへの欲求に影響を与えることを示しています。ベータエンドルフィンのレベルを低下させるオピエート受容体遮断薬を使用する際に、サルは手入れをしたいという欲求の高まりに応えました。対照的に、サルにモルヒネを投与すると、手入れをしたいという欲求が大幅に低下しました。ベータエンドルフィンは、脳脊髄液をサンプリングすることで測定できるオキシトシンとは異なり、動物種で測定するのが困難であり、したがって社会的行動とはそれほど強く関連し
糖質コルチコイド受容体
糖質コルチコイドは、副腎皮質で合成されるステロイドホルモンであり、コルチコステロイドのグループの一部です。糖質コルチコイドは免疫機能に関与しており、炎症を軽減するフィードバックシステムの一部です。さらに、糖質コルチコイドは糖代謝に関与している。マカクの研究は、社会的ストレスの増加が糖質コルチコイド耐性をもたらし、免疫機能をさらに阻害することを示しています。社会的グルーミングに参加したマカクは、ウイルス量のレベルの低下を示しました。これは、社会的ストレスのレベルの低下を示し、免疫機能と糖質コルチコイド感受性の増加をもたらします。さらに、1997年に発表された記事は、母親の手入れの増加が、新生児ラットの標的組織上の糖質コルチコイド受容体の比例した増加をもたらしたと結論付けました。新生児ラットに関する研究では、セロトニンと甲状腺刺激ホルモンの両方の濃度が変化したために、受容体の数が変化したことがわかった。受容体の数の増加は、コルチコステロイド分泌に対する負のフィードバックの量に影響を及ぼし、異常な生理的ストレス反応の望ましくない副作用を防ぐ可能性が身だしなみは糖質コルチコイド受容体の数を変える可能性があり、その結果免疫機能が高まる可能性が
研究はまた、社会的グルーミングにより多く参加するオスのヒヒは、より低い基礎コルチゾール濃度を示すことを示しました。
糞便糖質コルチコイド(fGC)は、ストレスに関連するホルモン代謝物であり、より強力で確立されたグルーミングネットワークを持つ雌のヒヒに低レベルで存在することが見られます。潜在的に乳児を殺すオスのヒヒがグループに移住すると、メスのfGCレベルが上昇するのが見られ、ストレスが高いことを示している。ただし、信頼性が高く確立されたグルーミングパートナーを持つ女性は、グルーミングネットワークが弱い女性よりもfGCの上昇が少なくなります。したがって、「友情」から受けた社会的支援は、ストレス管理においてヒヒを助けます。同様に、親しい「友人」が亡くなると、女性のfGCレベルも上昇することが見られます。ただし、これらの上昇するfGCレベルは、新しいグルーミングパートナーを形成する女性では減少し、亡くなった友人に取って代わります。
オピオイド
内因性オピオイドは、生物の脳で生成される化学分子であり、リラックス感、幸福感、痛みを和らげる働きをします。霊長類では、笑いと身だしなみが脳内のオピオイド放出を引き起こし、それが社会的絆を形成し維持すると考えられています。アカゲザルで実施された研究では、4〜10週齢の乳児を持つ授乳中の女性に、オピオイド受容体を遮断し内因性オピオイドの効果を阻害するオピオイド拮抗薬であるナロキソンを低用量で投与した。生理食塩水を与えられた対照の女性と比較して、ナロキソンの女性は彼らの乳児および彼らのグループの他のメンバーをあまり手入れしなかった。ナロキソンのメスはまた、若い母親の保護が弱いことが観察されました。これは、新しい母親の特徴ではありません。ナロキソン注射による社会的相互作用のこの低下は、オピオイド拮抗薬が社会的行動への母親の関与を妨げることを示唆しています-ここでは、社会的グルーミング。したがって、新しいアカゲザルの母親のオピオイドのレベルが高いと、社会的関与と「母親」の特性のレベルが高くなり、新生児の発達と学習を助けると仮定できます。
引用された研究に対する批判
とりわけ、社会的身だしなみに関する研究に関する主な批判は、それらのほとんどすべてが霊長目と霊長類自体の中の狭い範囲の種に焦点を合わせているということです。したがって、これは、社会的グルーミングの認知的または行動的基盤が何であるかについての包括的な考えを私たちに与えるものではなく、それのすべての効果(利益またはコスト)を完全に概説するものでもありません。さらに、十分に研究された種でさえ、社会的グルーミングに関連するすべてのデータを持っていない可能性が第二に、ほとんどの種のデータは、単一のグループのメンバーに基づいて導き出されます。社会環境条件に応じて行動が非常に柔軟な霊長類では、これは特別な課題となります。第三に、ほとんどの研究は観察的で短期的なものです。したがって、社会的身だしなみとフィットネスまたは配偶者選択の結果との直接的な関連は、長期的な直接または飼育下の研究のように直接研究することはできません。
も参照してください
掃除魚
清掃共生
コミュニティの結束 なめる 身繕い
社会的結束
参考文献
^ Henazi SP、Barrett L(1999年1月)。「女性霊長類への身づくろいの価値」。霊長類; 霊長類学ジャーナル。40(1):47–59。土井:10.1007 / BF02557701。PMID 23179531。S2CID 34106634。
^ Aureli F、Van Schaik CP、Van Hooff JA(1989)。「飼育下のカニクイザル(Macaca fascicularis)間の和解の機能的側面」。霊長類学のアメリカジャーナル。19(1):39–51。土井:10.1002 /ajp.1350190105。PMID 31964021。S2CID 86673865。
^ バートン、ロバート(1985年10月1日)。「霊長類のグルーミングサイトの好みとその機能的意味」。霊長類学の国際ジャーナル。6(5):519–532。土井:10.1007 / BF02735574。S2CID 37114535。
^ Aureli F、Waal FB(2000年1月1日)。自然な紛争解決。カリフォルニア大学出版。pp。193–224。ISBN
9780520223462。
^ Yee JR、Cavigelli SA、Delgado B、McClintock MK。「軽度のグループストレッサー中の青年期のラット間の相互所属は、乳腺腫瘍と寿命を予測します」。心身医学。70(9):1050–9。土井:10.1097 /psy.0b013e31818425fb。PMC 5767077。PMID 18842748。
^ Akinyi MY、Tung J、Jeneby M、Patel NB、Altmann J、Alberts SC。””Papio cynocephalus)””。動物の行動。85(3):559–568。土井:10.1016 /j.anbehav.2012.12.012。PMC 3961061。PMID 24659824。
^ ダンバー、ロビン(1991年1月1日)。「霊長類における社会的グルーミングの機能的重要性」。FoliaPrimatologica。57(3):121–131。土井:10.1159 / 000156574。
^ ボッチャ、マリア(1983年12月1日)。「アカゲザルの自己グルーミングとの直接比較による社会的グルーミングパターンの機能分析」。霊長類学の国際ジャーナル。4(4):399–418。土井:10.1007 / BF02735602。S2CID 24431777。
^ Seyfarth RM、Cheney DL(2011年7月5日)。「友情の進化の起源」。心理学の年次レビュー。63:153–77。土井:10.1146 / annurev-psych-120710-100337。PMID 21740224。
^ Schino G(2001年8月1日)。「女性霊長類間の身づくろい、競争および社会的地位:メタ分析」。動物の行動。62(2):265–271。土井:10.1006 /anbe.2001.1750。S2CID 53145237。
^ ウッドワード、アイリン。「霊長類の脳は、友人やVIPの顔に対して異なる反応を示します」。ニューサイエンティスト。
^ Roubová、ヴェロニカは、Konečná、Martina; Šmilauer、Petr; ウォールナー、バーナード(2015年2月10日)。「誰が花婿に、何のために?メスのバーバリーマカク(Macaca sylvanus)の毛づくろいのパターン」。PLOSONE。10(2):e0117298。Bibcode:2015PLoSO..1017298R。土井:10.1371 /journal.pone.0117298。PMC 4323340。PMID 25668722。
^ Carter G、Leffer L(2015年10月7日)。「コウモリの社会的グルーミング:吸血コウモリは例外的ですか?」。PLOSONE。10(10):e0138430。Bibcode:2015PLoSO..1038430C。土井:10.1371 /journal.pone.0138430。PMC 4596566。PMID 26445502。
^ Kumar、Rathina; カントール、マウリシオ; Senthilkumar、K。; Panneerselvam、Vimal; Kaliraj、P。; Marimuthu、G。(2017年1月1日)。「短い鼻のフルーツコウモリの間の社会的な手入れ」。行動。154:37–63。土井:10.1163 / 1568539X-00003410。
^ Yohe、Laurel R; ブランド、フィリップ。「化学感覚の進化生態学と感覚ドライブにおけるその役割」。現在の動物学。64(4):525–533。土井:10.1093 / cz / zoy048。ISSN 1674から5507まで。PMC 6084603。PMID 30108633。
^ Stopka、Pavel; ロマーナ、ストップコバ(2001年9月1日)。「ハーブフィールドマウスの条件付きグルーミング」。行動生態学。12(5):584–589。土井:10.1093 / beheco /12.5.584。
^ フェルプス、スティーブ; Ng、ウィング; Musolesi、Mirco; ラッセル、イヴァン(2018年9月11日)。「チンパンジーのグルーミングにおける正確なタイムマッチングは、少し遅れても起こりません」。PLOSONE。13(9):e0201810。Bibcode:2018PLoSO..1301810P。土井:10.1371 /journal.pone.0201810。PMC 6133271。PMID 30204753。
^ マーティン-オルダス、ジェマ; ハウン、ダニエル; コルメナレス、フェルナンド; ジョセップに電話してください。「時間を追跡する:類人猿におけるエピソードのような記憶の証拠」。動物の認知。13(2):331–340。土井:10.1007 / s10071-009-0282-4。ISSN 1435から9448まで。PMC 2822233。PMID 19784852。
^ Kutsukake N、Clutton-Brock TH(2010年2月1日)。「共同繁殖ミーアキャットにおける身づくろいと社会的関係の価値」。動物の行動。79(2):271–279。土井:10.1016 /j.anbehav.2009.10.014。S2CID 53203763。
^ Guan ZH、Huang B、Ning WH、Ni QY、Sun GZ、Jiang XL。「西部の黒い紋付きギボンの一夫多妻制の2つのグループにおける身づくろい行動の重要性:移民と居住者グループのメンバー間の社会的関係を理解するための含意」。霊長類学のアメリカジャーナル。75(12):1165–73。土井:10.1002 /ajp.22178。PMID 23843246。S2CID 205330388。
^ Carter G、Leffer L(2015年10月7日)。「コウモリの社会的グルーミング:吸血コウモリは例外的ですか?」。PLOSONE。10(10):e0138430。Bibcode:2015PLoSO..1038430C。土井:10.1371 /journal.pone.0138430。PMC 4596566。PMID 26445502。
^ 江、チー; Xia、Dong-Po; 王、西; 張、ダオ; 太陽、ビンファ; Li、Jin-Hua(2019)。「メスの中国チベットマカク(Macaca thibetana)における身づくろいと幼児の取り扱いの交換」。動物学研究。40(2):139–145。土井:10.24272 /j.issn.2095-8137.2018.049。PMC 6378558。PMID 29955029。
^ Surbeck、Martin; ホーマン、ゴットフリード(2014年12月18日)。「社会的嗜好は、男性のボノボの間での社会的グルーミングの短期間の交換に影響を与えます」。動物の認知。18(2):573–579。土井:10.1007 / s10071-014-0826-0。PMID 25519436。S2CID 16030429。
^ Silk JB、Alberts SC、Altmann J。「雌ヒヒの社会的絆は乳児の生存を高める」。科学。302(5648):1231–4。Bibcode:2003Sci … 302.1231S。土井:10.1126 /science.1088580。PMID 14615543。S2CID 24970809。
^ Archie EA、Tung J、Clark M、Altmann J、Alberts SC。「社会的所属は重要です:同性と異性の両方の関係は野生の雌のヒヒの生存を予測します」。議事録。生物科学。281(1793):20141261. DOI:10.1098 / rspb.2014.1261。PMC 4173677。PMID 25209936。
^ Sterck EH、Watts DP、Schaik CP(1997年11月1日)。「人間以外の霊長類における女性の社会的関係の進化」(PDF)。行動生態学および社会生物学。41(5):291–309。土井:10.1007 / s002650050390。hdl:1874/1183。ISSN 0340から5443まで。S2CID 25163826。
^ Silk JB、Alberts SC、Altmann J(2004年3月1日)。「ケニアのアンボセリにおける成体の雌ヒヒによる連合形成のパターン」。動物の行動。67(3):573–582。土井:10.1016 /j.anbehav.2003.07.001。S2CID 783193。
^ BerghänelA、Ostner J、SchröderU、SchülkeO(2011)。「ソーシャルボンドは、オスのバーバリーマカク、Macacasylvanusにおける将来の協力を予測します」。動物の行動。81(6):1109–1116。土井:10.1016 /j.anbehav.2011.02.009。S2CID 53144146。 ^ van Schaik CP、Kappeler PM(1997年11月)。「霊長類における子殺しのリスクと男女の関係の進化」。議事録。生物科学。264(1388):1687–94。Bibcode:1997RSPSB.264.1687V。土井:10.1098 /rspb.1997.0234。PMC 1688726。PMID 9404030。
^ グラハムB(2008年1月1日)。選択:進化のメカニズム。オックスフォード大学出版局。ISBN
978-0198569725。OCLC 781154368。
^ Okasha S(2013年1月1日)。Zalta EN(ed。)スタンフォード哲学百科事典(2013年秋版)。スタンフォード大学形而上学研究所。
^ ダーウィンC(1871)。人間の降下、および性との関係における選択。ジョンマレー。土井:10.5962 /bhl.title.2092。hdl:2027 / bc.ark:/ 13960 / t96696g2b。
^ Trivers RL(1971年3月1日)。「互恵的利他主義の進化」。生物学の四半期レビュー。46(1):35–57。土井:10.1086 / 406755。ISSN 0033から5770まで。S2CID 19027999。
^ Axelrod R、Hamilton WD(1981年3月)。「協力の進化」。科学。211(4489):1390–6。Bibcode:1981Sci … 211.1390A。土井:10.1126 /science.7466396。PMID 7466396。
^ 「種の起源:第VIII章。本能:本能に適用される自然淘汰の理論への異議:チャールズ・ダーウィン@クラシックリーダーによる中性および不妊虫」。www.classicreader.com 。
^ ライトS(1922年7月1日)。「近交係数と関係」。アメリカンナチュラリスト。56(645):330–338。土井:10.1086 / 279872。ISSN 0003から0147まで。S2CID 83865141。
^ スミスJM(1996年1月1日)。進化遺伝学。オックスフォード大学出版局。ISBN
978-0198542155。OCLC 954574132。
^ フランクR。「せっかちなトレーダーまたは偶発的な往復運動者?ヒヒの身づくろい交換の延長された時間経過の証拠」(PDF)。行動。146(8):1123–1135。doi:10.1163 / 156853909×406455 –Brill経由。
^ Barelli C、Reichard UH、Mundry R(2011年10月1日)。「グルーミングは野生のシロテテナガザル、Hylobates larの商品として使用されていますか?」。動物の行動。82(4):801–809。土井:10.1016 /j.anbehav.2011.07.012。S2CID 53190848。
^ 「霊長類の社会的グルーミング:個体発生」。www.reed.edu 。
^ モンドラゴン-Ceballosの、リカルド。Chiappa、Pilar; マヤゴイティア、リリアン; リー、フィリス(2010年7月1日)。「幼児のベニガオザルにおける社会的グルーミングの割り当ての学習における性差」。行動。147(9):1073〜1099。土井:10.1163 / 000579510X505436。hdl:1893/2990。
^ ワインバーグ、スーザンM。; Candland、Douglas K.(1981)。「ニホンザルの「石の手入れ」」。霊長類学のアメリカジャーナル。1(4):465–468。土井:10.1002 /ajp.1350010412。ISSN 1098年から2345年。PMID 31995914。S2CID 85138037。
^ Mcgrew、William; トゥティン、カロリン(1973年2月16日)。「歯科グルーミングにおけるチンパンジーツールの使用」。ネイチャー。241(5390):477–478。Bibcode:1973Natur.241..477M。土井:10.1038 / 241477a0。S2CID 4166467。
^ 「非人間霊長類による道具の使用|人類学の学術研究および訓練のためのセンター(CARTA)」。carta.anthropogeny.org 。
^ 「霊長類の認知| Scitableで科学を学ぶ」。www.nature.com 。
^ Moore D、Angel JE、Cheeseman IM、Robinson GE、Fahrbach SE(1995)。「高度に専門化された社会的グルーミングミツバチ(膜翅目:ミツバチ科)」。Journal of InsectBehavior。8(6):855–861。土井:10.1007 / BF02009512。S2CID 40236926。
^ Spruijt BM、van Hooff JA、Gispen WH(1992年7月)。「グルーミング行動の動物行動学と神経生物学」。生理学的レビュー。72(3):825–52。土井:10.1152 /physrev.1992.72.3.825。hdl:1874/3750。PMID 1320764。
^ Wilkinson GS(1986)。「ナミチスイコウモリDesmodusrotundusの社会的グルーミング」(PDF)。動物の行動。34(6):1880–1889。CiteSeerX 10.1.1.539.5104。土井:10.1016 / S0003-3472(86)80274-3。S2CID 11214563。
^ van Lawick-Goodall J(1968)。「ゴンベ渓流保護区における自由生活チンパンジーの行動」。動物の行動のモノグラフ。1:161–311。土井:10.1016 / s0066-1856(68)80003-2。
^ Waal F(1989)。霊長類間の平和構築。ケンブリッジ、マサチューセッツ州:ハーバード大学出版局。ISBN
9780674659209。
^ Smuts B、Cheney D、Seyfarth R、Wrangham R、Struhsaker T(1987)。霊長類の社会。シカゴ:シカゴ大学出版局。
^ カーター、ジェラルド; レファー、ローレン(2015年10月7日)。「コウモリの社会的グルーミング:吸血コウモリは例外的ですか?」。PLOSONE。10(10):e0138430。Bibcode:2015PLoSO..1038430C。土井:10.1371 /journal.pone.0138430。ISSN 1932から6203まで。PMC 4596566。PMID 26445502。
^ Schino G、Scucchi S、Maestripieri D、Turillazzi PG(1988)。「緊張緩和メカニズムとしてのグルーミング:行動的アプローチ」。霊長類学のアメリカジャーナル。16(1):43–50。土井:10.1002 /ajp.1350160106。PMID 31968881。S2CID 86566868。
^ Smuts BB(1985)。ヒヒのセックスと友情。テイラーアンドフランシスグループ。土井:10.4324 / 9781315129204。ISBN
978-1-315-12920-4。
^ 「霊長類の社会性と社会システム| Scitableで科学を学ぶ」。www.nature.com 。
^ 「霊長類の社会的な身づくろい:適応価値」。www.reed.edu 。
^ Schino、Gabriele; Aureli、Filippo(2008年11月1日)。「霊長類の身づくろいの往復運動におけるトレードオフ:行動の柔軟性と相関する進化のテスト」。リンネ協会の生物学ジャーナル。95(3):439–446。土井:10.1111 /j.1095-8312.2008.01067.x。ISSN 0024から4066まで。
^ Lehmann、Johannes。「霊長類におけるグループのサイズ、身づくろい、および社会的結束」(PDF)。動物の行動。74(6):1617–1629。土井:10.1016 /j.anbehav.2006.10.025。S2CID 14866172。
^ Kenny E、Birkhead TR、Green JP(2017年8月1日)。「鳥のアロプリーニングは、子孫の世話と長年にわたる安定したつがいをめぐる親の協力と関連しています」。行動生態学。28(4):1142–1148。土井:10.1093 / beheco / arx078。PMC 5873249。PMID 29622926。
^ Fraser ON、Bugnyar T。「ワタリガラスは慰めを示しますか?苦しんでいる他の人への反応」。PLOSONE。5(5):e10605。Bibcode:2010PLoSO … 510605F。土井:10.1371 /journal.pone.0010605。PMC 2868892。PMID 20485685。
^ Feh、C.、De Mazieres、J。好みの場所で手入れをすると、馬の心拍数が低下します。アニメーション。振る舞い。1993; 46:1191–1194。
^ Val-Laillet D、Guesdon V、Keyserlingk MA、PassilléAM、Rushen J(2009)。「牛のグルーミング:社会的嗜好、摂食移動および社会的支配の間の関係」。応用動物行動科学。116(2–4):141–149。土井:10.1016 /j.applanim.2008.08.005。
^ Laister S、Stockinger B、Regner A、Zenger K、Knierim U、Winckler C(2011)。「乳牛の社会的舐め—パフォーマーとレシーバーの心拍数への影響」。応用動物行動科学。130(3–4):81–90。土井:10.1016 /j.applanim.2010.12.003。
^ Kemp AH、Guastella AJ(2011年8月8日)。「人間の影響におけるオキシトシンの役割」。心理学の現在の方向性。20(4):222–231。土井:10.1177 / 0963721411417547。S2CID 145122328。
^ Uvnäs-Moberg K(1998年11月)。「オキシトシンは、前向きな社会的相互作用と感情の利益を仲介する可能性があります」。精神神経内分泌学。23(8):819–35。土井:10.1016 / S0306-4530(98)00056-0。PMID 9924739。S2CID 24789741。
^ Uvnäs-MobergK(1997年1月)。「社会的接触の生理学的および内分泌的影響」。ニューヨーク科学アカデミーの年報。807(1):146–63。Bibcode:1997NYASA.807..146U。土井:10.1111 /j.1749-6632.1997.tb51917.x。PMID 9071348。S2CID 46171612。
^ Dunbar RI。「人間と霊長類における接触の社会的役割:行動機能と神経生物学的メカニズム」。神経科学および生物行動学的レビュー。タッチ、温度、痛み/かゆみ、喜び。34(2):260–8。土井:10.1016 /j.neubiorev.2008.07.001。PMID 18662717。S2CID 30450770。
^ Keverne EB、Martensz ND、Tuite B(1989)。「サルの脳脊髄液中のベータエンドルフィン濃度は、身づくろいの関係に影響されます」。精神神経内分泌学。14(1–2):155–61。土井:10.1016 / 0306-4530(89)90065-6。PMID 2525263。S2CID 24350695。
^ ヘミングスHC(2013年1月1日)。麻酔のための生理学と薬理学:基礎と臨床応用:専門家による相談-オンラインと印刷。エルゼビア。ISBN
978-1437716795。OCLC 830351627。
^ Keverne EB、Martensz ND、Tuite B(1989年1月1日)。「サルの脳脊髄液中のベータエンドルフィン濃度は、身づくろいの関係に影響されます」。精神神経内分泌学。14(1–2):155–61。土井:10.1016 / 0306-4530(89)90065-6。PMID 2525263。S2CID 24350695。
^ Sapolsky RM、Romero LM、Munck AU。「糖質コルチコイドはストレス反応にどのように影響しますか?許容、抑制、刺激、および準備のアクションを統合します」。内分泌レビュー。21(1):55–89。土井:10.1210 /edrv.21.1.0389。PMID 10696570。
^ Capitanio JP、Mendoza SP、Lerche NW、Mason WA(1998年4月)。「社会的ストレスは、サル後天性免疫不全症候群において糖質コルチコイド調節の変化と生存期間の短縮をもたらします」。アメリカ合衆国科学アカデミー紀要。95(8):4714–9。Bibcode:1998PNAS … 95.4714C。土井:10.1073 /pnas.95.8.4714。PMC 22556。PMID 9539804。
^ Liu D、Diorio J、Tannenbaum B、Caldji C、Francis D、Freedman A、他 (1997年9月)。「母親のケア、海馬の糖質コルチコイド受容体、およびストレスに対する視床下部-下垂体-副腎の反応」。科学。277(5332):1659–62。土井:10.1126 /science.277.5332.1659。PMID 9287218。
^ Sapolsky RM(1997年9月)。「手入れの行き届いた子供の重要性」。科学。277(5332):1620–1。土井:10.1126 /science.277.5332.1620。PMID 9312858。S2CID 45145411。
^ Ray JC、Sapolsky RM(1992年1月1日)。「男性の社会的行動のスタイルとその内分泌は、上位の野生のヒヒの間で相関しています」。霊長類学のアメリカジャーナル。28(4):231–250。土井:10.1002 /ajp.1350280402。ISSN 1098年から2345年。PMID 31941207。S2CID 196609839。
^ Platt、Michael L。; Seyfarth、Robert M。; チェイニー、ドロシーL.(2016年2月5日)。「霊長類の脳における社会的認知への適応」。王立協会の哲学的取引B:生物科学。371(1687):20150096. DOI:10.1098 / rstb.2015.0096。ISSN 0962から8436まで。PMC 4745018。PMID 26729935。
^ Manninen、サンドラ; トゥオミネン、ラウリ; ダンバー、ロビンI。; Karjalainen、Tomi; ヒルボネン、ジュッシ; Arponen、Eveliina; ハリ、リーッタ; Jääskeläinen、Iiro P。; サムズ、ミッコ; ラウリ・ヌメンマー(2017年6月21日)。「社会的笑いは人間の内因性オピオイド放出を誘発する」。ジャーナルオブニューロサイエンス。37(25):6125–6131。土井:10.1523 /JNEUROSCI.0688-16.2017。ISSN 0270から6474まで。PMC 6596504。PMID 28536272。
^ マーテル、フランシスL。; ネビソン、クレアM。; レイメント、F。デビッド; シンプソン、マイケルJA; Keverne、Eric B.(1993年1月1日)。「オピオイド受容体遮断は、アカゲザルの母親の感情と社会的グルーミングを減少させます」。精神神経内分泌学。18(4):307–321。土井:10.1016 / 0306-4530(93)90027-I。ISSN 0306から4530まで。PMID 8391149。S2CID 11262534。
参考文献
Aureli F、van Schaik C、van Hooff J(1989)。「飼育下のカニクイザル(Macaca fascicularis)間の和解の機能的側面」。霊長類学のアメリカジャーナル。19(1):39–51。土井:10.1002 /ajp.1350190105。PMID 31964021。S2CID 86673865。
de Waal F(1989)。霊長類間の平和構築。ケンブリッジ、マサチューセッツ州:ハーバード大学出版局。ISBN 9780674659209。
Gumert MD。「マカクの交配市場における性の支払い」 (PDF)。動物の行動。74(6):1655–1667。土井:10.1016 /j.anbehav.2007.03.009。S2CID 9104008。
Keverne EB、Martensz ND、Tuite B(1989)。「サルの脳脊髄液中のベータエンドルフィン濃度は、身づくろいの関係に影響されます」。精神神経内分泌学。14(1–2):155–61。土井:10.1016 / 0306-4530(89)90065-6。PMID 2525263。S2CID 24350695。
木村R(1998)。「放し飼いの馬のバンドにおける相互グルーミングと好ましい仲間関係」。応用動物行動科学。59(4):265–276。土井:10.1016 / S0168-1591(97)00129-9。
リーPC(1987)。「同胞:未熟なベルベットモンキー間の協力と競争」。霊長類。28(1):47–59。土井:10.1007 / BF02382182。S2CID 21449948。
van Lawick-Goodall J(1968)。「ゴンベ渓流保護区における自由生活チンパンジーの行動」。動物の行動のモノグラフ。1:161–311。土井:10.1016 / S0066-1856(68)80003-2。
Moore D、Angel JE、Cheeseman IM、Robinson GE、Fahrbach SE(1995)。「高度に専門化された社会的グルーミングミツバチ(膜翅目:ミツバチ科)」。Journal of InsectBehavior。8(6):855–861。土井:10.1007 / BF02009512。S2CID 40236926。
サポルスキーRM(1997年9月)。「手入れの行き届いた子供の重要性」。科学。277(5332):1620–1。土井:10.1126 /science.277.5332.1620。PMID 9312858。S2CID 45145411。
Schino G、Scucchi S、Maestripieri D、Turillazzi PG(1988)。「緊張緩和メカニズムとしてのグルーミング:行動的アプローチ」。霊長類学のアメリカジャーナル。16(1):43–50。土井:10.1002 /ajp.1350160106。PMID 31968881。S2CID 86566868。
スマットB(1985)。ヒヒのセックスと友情。ニューヨーク:AldinePublications。ISBN 978-0-202-02027-3。
Smuts B、Cheney D、Seyfarth R、Wrangham R、Struhsaker T(1987)。霊長類の社会。シカゴ:シカゴ大学出版局。ISBN 9780226767161。
Spruijt BM、van Hooff JA、Gispen WH(1992年7月)。「グルーミング行動の動物行動学と神経生物学」。生理学的レビュー。72(3):825–52。土井:10.1152 /physrev.1992.72.3.825。hdl:1874/3750。PMID 1320764。
Vrontou S、Wong AM、Rau KK、Koerber HR、Anderson DJ。「invivoでの毛深い皮膚のマッサージのようなストロークを検出するC線維の遺伝的同定」。ネイチャー。493(7434):669–73。Bibcode:2013Natur.493..669V。土井:10.1038 / nature11810。PMC 3563425。PMID 23364746。
ウィルキンソンGS(1986)。「ナミチスイコウモリDesmodusrotundusの社会的グルーミング」 (PDF)。動物の行動。34(6):1880–1889。CiteSeerX 10.1.1.539.5104。土井:10.1016 / S0003-3472(86)80274-3。S2CID 11214563。
外部リンク
![]()
ウィクショナリーでのソーシャルグルーミングの辞書定義
コモンズでのソーシャルグルーミングに関連するメディア”