Somatosensory
「タッチ」はその他の使用法については、タッチを参照してください
「体性感覚システム」
体性感覚系は、の一部である感覚神経系のタッチの感覚に関連付けられているが、温度、身体の位置及び移動、及び痛みの感覚のための並列受容体および神経経路を含みます。感覚ニューロンと神経経路のこの複雑なシステムは、体の表面または内部の変化に反応します。感覚ニューロンの軸索(求心性神経線維として)は、さまざまな受容体細胞と接続するか、それに応答します。これらの感覚受容体細胞は、熱や侵害受容などのさまざまな刺激によって活性化されます、温度変化に関する情報を運ぶ熱受容体などの応答する感覚ニューロンに機能的な名前を付けます。他の受容体タイプには、機械受容器、化学受容器、および侵害受容器が含まれ、これらは感覚神経に沿って脊髄に信号を送り、そこで信号は他の感覚ニューロンによって処理され、その後、さらなる処理のために脳に中継されます。感覚受容体は、皮膚、上皮組織、筋肉、骨や関節、内臓、心臓血管系など、全身に見られます。
タッチは情報を受け取るための重要な手段です。この写真は、視覚障害者の階段を識別する触覚マーキングを示しています。
体性感覚は、時にはと呼ばれる体感感覚、 somesthesisが含まれていることを理解して触覚を、固有感覚(位置及び動きの感覚)、[] (使用に依存)と触覚感覚。
脳内の体表面のマッピングはソマトトピーと呼ばれます。皮質では、それは皮質ホムンクルスとも呼ばれます。ただし、この脳表面(「皮質」)マップは不変ではありません。脳卒中やけがに反応して劇的な変化が起こる可能性が
コンテンツ
1 システム概要
1.1 感覚受容器 1.2 体性感覚皮質
2 構造
3 一般的な体性感覚経路
3.1 触覚フィードバック 3.2 バランス 3.3 ファインタッチとラフタッチ 3.43.4 社会的タッチの神経処理 3.5 個人差
4 臨床的な意義
5 社会と文化
6 も参照してください
7 ノート
8 参考文献
9 参考文献
10 外部リンク
システム概要
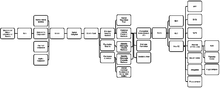
この図は、(特に明記されていない限り)線形に、人間の脳内の関連するエンドポイントへの接触を可能にするすべての既知の構造の投影を追跡します。
感覚受容器
皮膚の4つの機械受容器は、それぞれ異なる刺激に短期間または長期間反応します。
メルケル細胞の神経終末は、基底表皮と毛包に見られます。それらは、低振動(5〜15 Hz)および形状やエッジなどの深い静的タッチに反応し ます。受容野が小さいため(非常に詳細な情報)、指先などの領域で最もよく使用されます。それらは覆われていない(殻から取り出されている)ため、長期間にわたって圧力に反応します。
触覚小体は中程度の振動(10〜50 Hz)と軽い接触に反応します。それらは真皮乳頭にあります; それらの反応性のために、それらは主に指先と唇にメルケル盤とは異なり、それらは迅速な活動電位で反応します。彼らは点字を読み、穏やかな刺激を感じる能力に責任が
パチニ小体は、全体的なタッチを決定し、粗い物質と柔らかい物質を区別します。それらは、特に250 Hz付近(最大センチメートル離れた場所)の振動に対して、迅速な活動電位で反応します。それらは振動に最も敏感であり、大きな受容野を持っています。パチニ小体は突然の刺激にのみ反応するため、常に形状を圧縮している衣服のような圧力はすぐに無視されます。それらはまた、ハンドヘルドツールのタッチ感覚の位置を検出することにも関係しています。
ルフィニ小体はゆっくりと反応し、持続的な皮膚のストレッチに反応します。それらは物体の滑りの感覚に責任があり、運動感覚と指の位置と動きの制御に大きな役割を果たします。メルケルと球根細胞-反応が遅い-は有髄です; 残りの-高速応答-はそうではありません。これらの受容体はすべて、その形状を押しつぶして活動電位を引き起こす圧力によって活性化されます。
体性感覚皮質
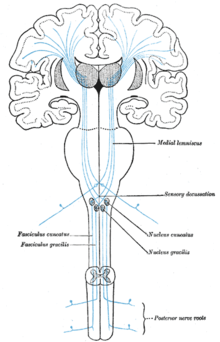
グレイの解剖学、図759:感覚路、体性感覚視床を通り、S1(ブロードマンの脳地図3、1、および2)、S2、およびBA7への脊髄の経路(青)を示しています

グレイの解剖学、図717:島皮質(この図で島とマークされている)に隣接し、S1、S2、およびBA7に
隣接する経路を示す詳細
中心後回は、一次体性感覚皮質(ブロードマン領域 3,2及び1をまとめS1と記します)。
BA3は、視床から最も密度の高い投影を受け取ります。BA3aは、隣接する身体部分の相対的な位置の感覚と、運動中に使用される努力の量に関係しています。BA3bは体性感覚情報の配信を担当し、テクスチャ情報をBA1に投影し、形状とサイズの情報をBA2に投影します。
領域S2(二次体性感覚野)は、領域S2と頭頂腹側領域に分かれています。エリアS2は特定の触覚に関与しているため、扁桃体および海馬と一体的にリンクして、記憶をエンコードおよび強化します。
頭頂腹側領域は、運動前野および体性感覚記憶ハブ、BA5への体性感覚リレーです。
BA5は、地形的に組織化された体細胞記憶野および連合野です。
BA1はテクスチャ情報を処理し、BA2はサイズと形状の情報を処理します。
エリアS2は、軽いタッチ、痛み、内臓感覚、および触覚的注意を処理します。
S1は残りの情報(粗いタッチ、痛み、体温)を処理します。
BA7は、視覚情報と固有受容情報を統合して、空間内のオブジェクトを特定します。
島皮質(insula)は、身体の所有権、身体の自己認識、および知覚の意味での役割を果たしています。島はまた、官能的な接触、痛み、体温、かゆみ、および局所的な酸素状態に関する情報を伝達する役割も果たします。島は高度に接続されたリレーであるため、多くの機能に関与しています。
構造
体性感覚システムは、脊椎動物の体のすべての主要部分に広がっています。それは、中枢神経系内のより深いニューロンまで、末梢(例えば、皮膚、筋肉、器官)の感覚受容体と感覚ニューロンの両方で構成されています。
一般的な体性感覚経路
参照:後
柱-内側レムニスカス経路
すべての求心性接触/振動情報は、薄筋(T7以下)または楔状束(T6以上)を介して後部(背側)柱-内側レムニスカス経路を介して脊髄を上昇します。楔状束は、脊髄灰白質を介して間接的に蝸牛神経核に信号を送信します。この情報は、知覚される音が単なる絨毛ノイズ/刺激であるかどうかを判断するために使用されます。すべての繊維は延髄で交差します(左が右になります)。
体性感覚経路には通常、3つのニューロンがあります: 1次、2次、および3次。
一次ニューロンのタイプでpseudounipolarニューロンと常に有するセル本体に後根神経節の脊髄神経周と軸索タッチ神経支配メカノと二次ニューロン上の中央軸索シナプスを形成します。体性感覚経路が頸神経で覆われていない頭または首の部分にある場合、一次ニューロンは三叉神経節または他の感覚脳神経節になります)。
二次ニューロンは、その有する細胞体を、脊髄または脳幹のいずれか。このニューロンの上行軸索は、脊髄または脳幹のいずれかで反対側に交差(議論)します。
触覚や特定の種類の痛みの場合、三次ニューロンは視床の腹側後核に細胞体を持ち、一次体性感覚野(またはS1)の頭頂葉の中心後回で終わります。

タッチは、多くの異なる生理学的反応を引き起こす可能性がここでは、赤ちゃんが姉にくすぐられて笑ってい 目の網膜に見られるものと同様の光受容体は、潜在的に損傷を与える紫外線(特に紫外線A)を検出し、メラノサイトによるメラニン生成の増加を誘発します。したがって、日焼けは、紫外線によって引き起こされるDNA損傷および日焼け(紫外線Bによって引き起こされるDNA損傷)から皮膚を迅速に保護する可能性がしかし、これが保護を提供するかどうかは議論の余地がなぜなら、このプロセスによって放出されるメラニンの量は、紫外線B放射によって引き起こされるDNA損傷に応答して放出される量と比較して控えめだからです。
触覚フィードバック
固有受容感覚からの触覚フィードバックは、皮膚、筋肉、関節の固有受容器に由来します。
バランス
平衡感覚の受容体は、耳の前庭系にあります(頭の3次元の向き、および推測によると、体の残りの部分)。バランスは、固有受容感覚(頭に対する体の残りの部分の相対的な位置を感知する)によって供給される運動感覚反射によっても媒介されます。さらに、固有受容感覚は、身体の機械的反射への入力として、視覚系(身体に対するそれらの物体の位置の確認を提供する)によって感知される物体の位置を推定します。
ファインタッチとラフタッチ 参照: 2点識別
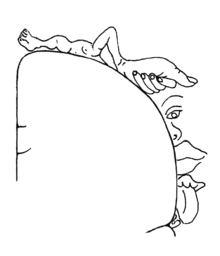
皮質ホムンクルス、脳の体性感覚領域のマップは、によって考案された
ワイルダー・ペンフィールド。
ファインタッチ(または識別タッチ)は、被験者がタッチを感知して位置を特定できるようにする感覚モダリティです。ローカリゼーションが不可能なタッチの形式は、粗いタッチとして知られています。後部列内側毛帯経路はに細かいタッチ情報の送信を担当する経路である大脳皮質の脳の。
粗いタッチ(または非識別的タッチ)は、被験者が触れられた場所を特定することなく(「ファインタッチ」とは対照的に)、何かが触れられたことを被験者が感知できるようにする感覚モダリティです。その繊維は、後柱で運ばれる細かいタッチとは異なり、脊髄視床路で運ばれます。ファインタッチは通常、粗タッチと並行して機能するため、ファインタッチを運ぶ繊維(後柱-内側毛帯経路)が破壊されるまで、人はタッチを局所化することができます。その後、被験者はタッチを感じますが、タッチされた場所を特定することはできません。
社会的タッチの神経処理
体性感覚皮質は、全身の受容体から入ってくる感覚情報をエンコードします。感情的なタッチは、感情的な反応を誘発する一種の感覚情報であり、通常、物理的な人間のタッチなど、本質的に社会的です。このタイプの情報は、実際には他の感覚情報とは異なる方法でコード化されています。感情的なタッチの強度は、一次体性感覚野でまだエンコードされており、身体的能力の欠如とは対照的に、愛する人の社会的タッチによって引き起こされるアドレナリンの増加によって例示されるように、視覚と音によって引き起こされる感情と同様の方法で処理されます愛していない人に触れること。
一方、感情的なタッチに関連する心地よさの感覚は、一次体性感覚野よりも前帯状皮質を活性化します。機能的磁気共鳴画像法(fMRI)データは、前帯状皮質と前頭前野の血中酸素濃度コントラスト(BOLD)信号の増加が、感情的なタッチの心地よさのスコアと高度に相関していることを示しています。阻害性経頭蓋磁気刺激の一次体性感覚皮質の(TMS)は、情動タッチ強度の知覚ではなく、感情タッチ快を阻害します。したがって、S1は、社会的に感情的なタッチの心地よさの処理には直接関与しませんが、タッチの位置と強度を区別する役割を果たします。
個人差
さまざまな研究が、細かい接触の意味での個人間の違いの原因を測定し、調査してきました。よく研究されている領域の1つは、受動的な触覚の空間的鋭敏さです。これは、静止した皮膚に押し付けられたオブジェクトの微細な空間的詳細を解決する機能です。受動的な触覚の空間的鋭敏さを測定するためにさまざまな方法が使用されてきましたが、おそらく最も厳密なのは格子配向タスクです。このタスクでは、被験者は2つの異なる方向で提示された溝付き表面の方向を特定します。これは手動または自動装置で適用できます。多くの研究は、年齢とともに受動的な触覚の空間的鋭敏さの低下を示しています。 この低下の理由は不明ですが、通常の老化中の触覚受容体の喪失が含まれる可能性が驚くべきことに、人差し指の受動的な触覚の空間的鋭敏さは、人差し指の指先が小さい成人の間で優れています。指のサイズのこの効果は、男性と比較して、平均して、女性のより良い受動的な触覚の空間的鋭敏さの根底にあることが示されています。低周波振動を検出する機械受容器の一種である触覚小体の密度は、小さい指ほど大きくなります。同じことがメルケル細胞にも当てはまる可能性がメルケル細胞は、微細な空間視力にとって重要な静的なくぼみを検出します。同じ年齢の子供たちの間では、指が小さい子供たちも触覚の鋭敏さが優れている傾向が多くの研究は、受動的触覚空間視力が同じ年齢の視力のある個人と比較して視覚障害者の間で強化されることを示しています おそらく大脳のクロスモーダル可塑性のため視覚障害者の皮質。おそらくまた、皮質の可塑性のために、生まれてから盲目である個人は、視力のある人々よりも迅速に触覚情報を統合すると報告されています。
臨床的な意義
体性感覚障害
体性感覚障害は、体性感覚系の末梢神経が関与する末梢神経障害によって引き起こされる可能性がこれはしびれや知覚異常として現れることが
社会と文化
ハプティックテクノロジーと
ハプティックコミュニケーション
触覚技術は、仮想環境と実環境でタッチ感覚を提供できます。言語療法の分野では、触覚フィードバックを使用して言語障害を治療することができます。
も参照してください
アロキリア
細胞シグナル伝達
ゴルジ腱器官
触覚コミュニケーション
触覚知覚
筋紡錘
分子細胞認知
幻肢
肉体的親密さ
感覚マップ
特殊感覚
縁上回
触覚錯覚
Vibratese、タッチによるコミュニケーションの方法
触覚イメージング
ノート
^ ピエゾチャネル受容体は、圧力、触覚、固有受容感覚の知覚において重要な役割を果たします(ピエゾ2受容体)。
^ TRPV1およびTRPM8受容体は、暑さと寒さの知覚において重要な役割を果たします。
参考文献
^ シャーマン、カール(2019年8月12日)。「感覚:体性感覚システム」。ダナ財団。ニューヨーク。
^ カロリンスカ研究所でのノーベル会議(2021年10月4日)プレスリリース:ノーベル生理学・医学賞2021ノーベル生理学・医学賞2021:デビッド・ジュリアス、アーデム・パタポウティアン
^ O. Franzen、R。Johansson、L。Terenius(1996)体性感覚皮質の感覚と神経生物学
^ Robles-De-La-Torre G(2006)。「仮想および実環境における触覚の重要性」(PDF)。IEEEマルチメディア。13(3):24–30。土井:10.1109 /MMUL.2006.69。S2CID 16153497。
^ シマ、リチャード(2019年12月23日)。「脳は体を超えたタッチを感知する」。サイエンティフィックアメリカン。
^ パレ、ミシェル、キャサリンベーツ。「人間の人差し指パッドにおける推定ルフィニ小体の不足。」ワイリーオンラインライブラリ。2003年2月10日。Web。2016年3月27日。
^ Scheibert J、Leurent S、Prevost A、DebrégeasG。「生体模倣センサーで精査された触覚情報のコーディングにおける指紋の役割」。科学。323(5920):1503–6。arXiv:0911.4885。Bibcode:2009Sci … 323.1503S。土井:10.1126 /science.1166467。PMID 19179493。S2CID 14459552。
^ Biswas A、Manivannan M、Srinivasan MA(2015)。「振動触覚感度閾値:パチニ小体の非線形確率的メカノトランスダクションモデル」。HapticsでのIEEEトランザクション。8(1):102–13。土井:10.1109 /TOH.2014.2369422。PMID 25398183。S2CID 15326972。
^ パレ、ミシェル、ロバートエルデ。「改訂されたマイスナー小体:侵害受容器の免疫化学的特性を備えた多求心性機械受容器。」JNeurosci。2001年9月15日。Web。2016年3月27日。
^ Hashim IH、熊本S、竹村K、前野T、奥田S、森Y。「人間の触覚メカニズムに触発された多層構造の触覚評価フィードバックシステム」。センサー(スイス、バーゼル)。17(11):2601 Bibcode:2017Senso..17.2601H。土井:10.3390 / s17112601。PMC 5712818。PMID 29137128。
^ Buccino G、Binkofski F、Fink GR、Fadiga L、Fogassi L、Gallese V、Seitz RJ、Zilles K、Rizzolatti G、Freund HJ。「行動観察は、運動前野と頭頂葉を体性的に活性化する:fMRI研究」。神経科学のヨーロッパジャーナル。13(2):400–4。土井:10.1111 /j.1460-9568.2001.01385.x。PMID 11168545。S2CID 107700。
^ Seelke AM、Padberg JJ、Disbrow E、Purnell SM、Recanzone G、Krubitzer L。「マカクザルのブロードマンの脳地図5内の地形図」。大脳皮質。22(8):1834–50。土井:10.1093 / cercor / bhr257。PMC 3388892。PMID 21955920。
^ Geyer S、Schleicher A、Zilles K(1999年7月)。「ヒト一次体性感覚野の領域3a、3b、および1」。NeuroImage。10(1):63–83。土井:10.1006 /nimg.1999.0440。PMID 10385582。S2CID 22498933。
^ Disbrow E。「マカクザルの頭頂腹側領域(PV)と第2体性感覚野(S2)の視床皮質接続」。視床および関連システム。1(4):289–302。土井:10.1016 / S1472-9288(02)00003-1。
^ サラディンKS。解剖学および生理学第3版。2004年。ニューヨークのマグロウヒル。
^ Zukerman、ウェンディ。「肌は日光から守るために光を見る」。newscientist.com。ニューサイエンティスト。
^ Proske U、サウスカロライナ州ガンデビア。「固有受容感覚:体の形、体の位置と動き、および筋力の信号伝達におけるそれらの役割」。生理学的レビュー。92(4):1651–97。土井:10.1152 /physrev.00048.2011。PMID 23073629。
^ Proske U、サウスカロライナ州ガンデビア。「運動感覚」。生理学ジャーナル。587(Pt 17):4139–46。土井:10.1113 /jphysiol.2009.175372。PMC 2754351。PMID 19581378。
^ Case LK、Laubacher CM、Olausson H、Wang B、Spagnolo PA、Bushnell MC。「人間の一次体性感覚野における触覚強度の符号化は心地よさではない」。ジャーナルオブニューロサイエンス。36(21):5850–60。土井:10.1523 /JNEUROSCI.1130-15.2016。PMC 4879201。PMID 27225773。
^ Van Boven、RW; ジョンソン、KO(1994年12月1日)。「人間の触覚空間分解能の限界:唇、舌、指での格子方向の識別」。神経学。44(12):2361–6。土井:10.1212 /WNL.44.12.2361。PMID 7991127。S2CID 32255147。
^ Craig JC(1999)。「触覚の空間的鋭敏さの尺度としての格子配向」。体性感覚および運動研究。16(3):197–206。土井:10.1080 / 08990229970456。PMID 10527368。
^ Goldreich D、Wong M、Peters RM、Kanics IM。「触覚自動パッシブフィンガースティミュレーター(TAPS)」。Journal of Visualized Experiments(28)。土井:10.3791 / 1374。PMC 2726582。PMID 19578327。
^ Stevens JC、Alvarez-Reeves M、Dipietro L、Mack GW、Green BG(2003)。「加齢における触覚の鋭敏さの低下:身体部位、血流、および喫煙と身体活動の生涯の習慣の研究」。体性感覚および運動研究。20(3–4):271–9。土井:10.1080 / 08990220310001622997。PMID 14675966。S2CID 19729552。
^ Manning H、Tremblay F(2006)。「指先での触覚パターン認識の年齢差」。体性感覚および運動研究。23(3–4):147–55。土井:10.1080 / 08990220601093460。PMID 17178550。S2CID 24407285。
^ Goldreich D、Kanics IM。「視覚障害では触覚が向上します」。ジャーナルオブニューロサイエンス。23(8):3439–45。土井:10.1523 /jneurosci.23-08-03439.2003。PMC 6742312。PMID 12716952。
^ Peters RM、Hackeman E、Goldreich D。「指小辞の数字は繊細な詳細を識別します:指先のサイズと触覚の空間的鋭敏さの性差」。ジャーナルオブニューロサイエンス。29(50):15756–61。土井:10.1523 /JNEUROSCI.3684-09.2009。PMC 3849661。PMID 20016091。
^ Dillon YK、Haynes J、Henneberg M。「マイスナー小体の数と皮膚グリフ文字および指のサイズとの関係」。解剖学ジャーナル。199(Pt 5):577–84。土井:10.1046 /j.1469-7580.2001.19950577.x。PMC 1468368。PMID 11760888。
^ Peters RM、Goldreich D(2013)。「小児期の触覚空間視力:年齢と指先の大きさの影響」。PLOSONE。8(12):e84650。Bibcode:2013PLoSO … 884650P。土井:10.1371 /journal.pone.0084650。PMC 3891499。PMID 24454612。
^ スティーブンス、ジョセフC。; Foulke、Emerson; パターソン、マシューQ.(1996)。「長期的な失明における触覚の鋭敏さ、老化、および点字の読書」。Journal of Experimental Psychology:Applied。2(2):91–106。土井:10.1037 /1076-898X.2.2.91。
^ Van Boven RW、Hamilton RH、Kauffman T、Keenan JP、Pascual-Leone A。「ブラインド点字リーダーの触覚空間分解能」。神経学。54(12):2230–6。土井:10.1212 /wnl.54.12.2230。PMID 10881245。S2CID 12053536。
^ Goldreich D、Kanics IM。「触覚格子検出タスクにおける視覚障害者と視力のある人間のパフォーマンス」。知覚と精神物理学。68(8):1363–71。土井:10.3758 / bf03193735。PMID 17378422。
^ Wong M、Gnanakumaran V、Goldreich D。「視覚障害における触覚の空間的視力の向上:経験に依存するメカニズムの証拠」。ジャーナルオブニューロサイエンス。31(19):7028–37。土井:10.1523 /JNEUROSCI.6461-10.2011。PMC 6703211。PMID 21562264。
^ Bhattacharjee A、Ye AJ、Lisak JA、Vargas MG、Goldreich D。「振動触覚マスキング実験は、先天的に盲目の点字リーダーにおける加速された体性感覚処理を明らかにします」。ジャーナルオブニューロサイエンス。30(43):14288–98。土井:10.1523 /JNEUROSCI.1447-10.2010。PMC 3449316。PMID 20980584。
^ GabrielRobles-De-La-Torre。「InternationalSocietyfor Haptics:Haptic technology、animateddescription」。Isfh.org。アーカイブされたオリジナルの2010年3月7日に。
参考文献
Boron WF、Boulpaep EL(2003)。医学生理学。サンダース。pp。352–358。ISBN 0-7216-3256-4。
フラナガン、JR、レダーマン、SJ神経生物学:隆起と穴を感じる、ニュースとビュー、ネイチャー、2001年7月26日; 412(6845):389-91。
Hayward V、Astley OR、Cruz-Hernandez M、Grant D、Robles-De-La-Torre G(2004)。「触覚インターフェースとデバイス」 (PDF)。センサーレビュー。24(1):16–29。土井:10.1108 / 02602280410515770。
Purves、Dale(2012)。神経科学、第5版。マサチューセッツ州サンダーランド:Sinauer Associates、Inc。pp。202–203。ISBN 978-0-87893-695-3。
Robles-De-La-Torre G、Hayward V。「力は、アクティブなタッチを通じて形状の知覚においてオブジェクトの幾何学を克服することができます」 (PDF)。ネイチャー。412(6845):445–8。Bibcode:2001Natur.412..445R。土井:10.1038 / 35086588。PMID 11473320。S2CID 4413295。
Robles-De-La-Torre、G(2006)。「仮想および実環境における触覚の重要性」 (PDF)。IEEEマルチメディア。13(3):24–30。土井:10.1109 /mmul.2006.69。S2CID 16153497。
Grunwald、M。(Ed。)Human Haptic Perception – Basics andApplications。ボストン/バーゼル/ベルリン:ビルクホイザー、2008年、
ISBN 978-3-7643-7611-6
タッチ スカラーペディアエキスパート記事の百科事典
外部リンク
コモンズの体性感覚に関連するメディア
タッチの解剖学。BBCラジオ4による事実上のドキュメンタリーシリーズ。”