Tc1/mariner
は、ドメイン、構造
眠れる森の美女トランスポゾンシステムには、いくつかの素晴らしい数字が
Tc1 / marinerは、散在するリピート DNA(クラスII)トランスポゾンのクラスおよびスーパーファミリーです。このクラスの要素は、人間を含むすべての動物に見られます。それらは原生生物やバクテリアにも見られます。
このクラスは、最もよく研究されている2つのメンバー、CaenorhabditiselegansのTc1トランスポゾンとDrosophilaのマリナートランスポゾンにちなんで名付けられました。
コンテンツ
1 構造
1.1 トランスポザーゼ
2 サブグループ
2.1 Tc1 2.2 マリナー 2.3 他の家族
3 トリビア
4 も参照してください
5 参考文献
構造

A。DNAトランスポゾンの構造(マリナー型)。
B。転置のメカニズム
トランスポゾンは、2つの末端逆方向反復配列(TIR)に隣接するトランスポザーゼ遺伝子で構成されています。2つの短いタンデムサイト重複(TSD)がインサートの両側に存在します。転位は、2つのトランスポザーゼがTIR配列を認識して結合し、結合してDNA二本鎖切断を促進するときに起こります。次に、DNA-トランスポザーゼ複合体は、そのDNAカーゴをゲノムの他の場所の特定のDNAモチーフに挿入し、統合時に短いTSDを作成します。 IS630 / Tc1 / marinerシステムでは、使用されるモチーフは「TA」ジヌクレオチドであり、挿入後に両端で複製されます。
トランスポザーゼ遺伝子がトランスポゾンによって運ばれないとき、それは今や遺伝子が動き回るために他の場所で発現されることを必要とするという点で非自律的になります。
トランスポザーゼ
トランスポザーゼ、タイプ1(部分的なDDEドメイン)
識別子
シンボル
トランスポザーゼ_1Pfam F01359
Pfam氏族L0219 InterPro PR001888 CATH u7b
利用可能なタンパク質構造:
Pfam
構造/ ECOD PDB RCSB PDB ; PDBe ; PDBj PDBsum 構造の概要
Mos1トランスポザーゼのHTHドメイン
識別子
シンボルTH_48 Pfam F17906
Pfam氏族L0123 InterPro PR041426 CATH u7b
利用可能なタンパク質構造:
Pfam
構造/ ECOD PDB RCSB PDB ; PDBe ; PDBj PDBsum 構造の概要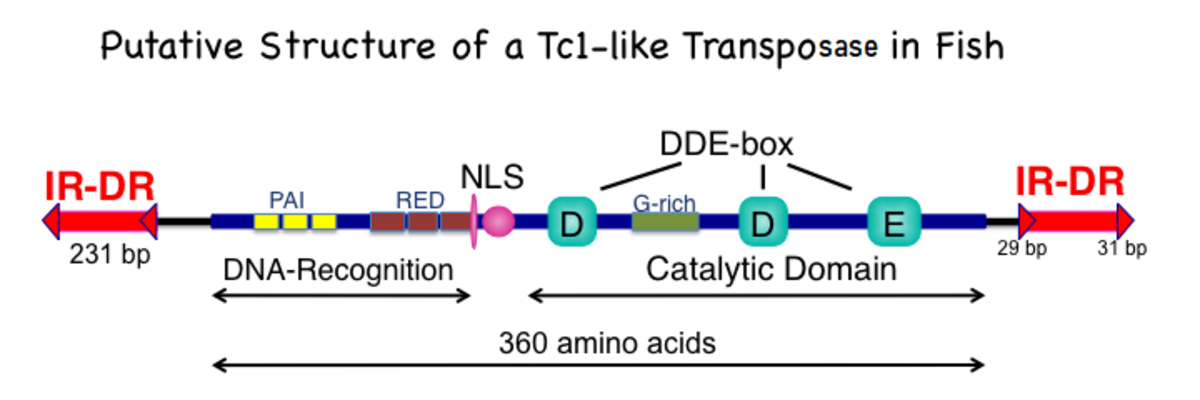
SBトランスポザーゼの構造的特徴。
360アミノ酸のポリペプチドには、3つの主要なサブドメインがトランスポゾンのミラーリングされたIR / DR配列のDR配列、核局在化配列(NLS)、およびDDEへの結合に関与するアミノ末端DNA認識ドメインです。転位を構成する一連のカットアンドペースト反応を触媒するドメイン。DNA認識ドメインには、DNAに結合でき、いくつかの転写因子に見られるさまざまなモチーフに関連する2つのペアのボックス配列が2つのペアのボックスにはPAIとREDのラベルが付いており、どちらもDNA結合ドメインに共通のヘリックスターンヘリックスモチーフを持っています。触媒ドメインには、多くのトランスポザーゼおよびリコンビナーゼ酵素に見られる特徴的なDDE(場合によってはDDD)アミノ酸がさらに、グリシン(G)アミノ酸が非常に豊富な領域が
タンパク質のマルチドメインの性質を考慮して、転写酵素のスーパーファミリーのいくつかの署名がさまざまなドメインデータベースで提供されています。さらに、各ドメインは、ボックスの「PAI」の半分のPF17906 / PF01710 / PF11427など、複数のエントリで表されることがよくREDボックスも同様にダイビング(PF08279 / PF13412 / PF01498など)であり、DNA認識用の翼のあるHTH形式であることがよく
サブグループ
Tc1 / marinerスーパーファミリーは、一般に、そのトランスポザーゼの触媒ドメインによって細分されます。通常、DDE(Asp-Asp-Glu)またはDDD触媒トライアドを使用します。
Tc1
Tc1(DD34E)は、Caenorhabditiselegansで活性のあるトランスポゾンです。 ヒトにはTc1のようなトランスポゾンもあり、すべて不活性です。Tc1のような要素は、いくつかの魚種や両生類を含む他の下等脊椎動物に存在します。
でC.エレガンスは、1610塩基対長の配列です。実験によると、この要素はヒト細胞で「ジャンプ」し、必要なタンパク質はトランスポザーゼだけです。
このファミリーのもう1つの例は、C。elegansに見られるトランスポゾンであるTc3です。
マリナー
マリナー(DD34D)の要素は、人間を含む複数の種に見られます。 マリナートランスポゾンは、最初にヤコブソンとHARTLによって発見されたショウジョウバエ1986年 、グループの分類は、1993年に出版されたmauritianaに昆虫におけるような配列を分割した、セクロピア、ミツバチ、irritans、とcapitata亜科、それらが発見された昆虫の種類の後。分類は他の種にも及ぶ。
この転移因子は、多くの種で水平感染するというその驚くべき能力で知られています。 ヒトゲノムには、260万塩基対からなる推定14,000コピーのマリナーが存在します。動物以外の最初のマリナー要素トランスポゾンは、膣トリコモナスで発見されました。
ヒトマリナーのようなトランスポゾンは、Hsmar1(セクロピア)とHsmar2(イリタン)のサブファミリーに分けられます。どちらのタイプも不活性ですが、SETMAR遺伝子にあるHsmar1の1つのコピーは、ヒストン修飾タンパク質にDNA結合を提供するため、選択中です。 Hsmar2は、化石シーケンスから複数回再構築されています。
Mos1(モザイク要素用)はDrosophilamauritianaで発見されました。 Himar1エレメントは、ノサシバエHaematobia irritansから分離されており、大腸菌の遺伝的ツールとして使用できます。
他の家族
rosa(DD41D)ファミリーは、Ceratitisrosaに見られるファミリーです。 Pogo / Fot1(DDxD)は、このスーパーファミリーのさらに別のファミリーであり、xは可変長を示します。IS630、中に可動要素赤痢ソンネは、また、このスーパーファミリーに属します。
トライアド間で長さが異なるいくつかの新しいファミリが報告されています。
ヒトではティガーとしても知られているポゴは、ヒトと酵母によって同様にCENPB遺伝子に家畜化されています。ポゴの他の人間の家畜化には、TIGD1、TIGD2、TIGD3、TIGD4、TIGD5、TIGD6、TIGD7、JRK、JRKL、POGK、およびPOGZが含まれます。
トリビア
マリナートランスポゾンの特徴は、ボブマーによるSF小説マリナープロジェクトに影響を与えました。
も参照してください
眠れる森の美女トランスポゾンシステム
PiggyBacトランスポゾンシステム
参考文献
^ Plasterk、Ronald HA; Izsvák、Zsuzsanna; Ivics、Zoltán(1999)。「常駐エイリアン:転移因子のTc1 / marinerスーパーファミリー」。遺伝学の動向。15(8):326–32。土井:10.1016 / S0168-9525(99)01777-1。PMID 10431195。
^ ロバートソン、HM(1995)。「動物におけるトランスポゾンのTc1-marinerスーパーファミリー」。J.昆虫の生理。41(2):99–105。土井:10.1016 / 0022-1910(94)00082-r。
^ Bradic、Martina; ウォーリング、サリーD; 低、ビビアン; カールトン、ジェーンM(2014)。「Tc1 / mariner転移因子ファミリーは原生生物Trichomonasvaginalisの遺伝的変異と遺伝子発現を形作る」。モバイルDNA。5:12。DOI:10.1186 / 1759-8753-5-12。PMC 4021607。PMID 24834134。 ^ Capy、Pierre; ランギン、ティエリー; ヒゲ、ドミニク; マウラー、パトリシア; バジン、クロード(1997)。「LTRレトロトランスポゾンとクラスIIエレメントトランスポザーゼのインテグラーゼには共通の祖先がありますか?」Genetica。100(1/3):63–72。土井:10.1023 / A:1018300721953。PMID 9440259。S2CID 24866580。 ^ Walter、Marius(2016)。DNAメチル化の動的喪失時のトランスポゾン調節(論文)。ピエールエマリーキュリー大学。土井:10.13140 /rg.2.2.18747.21286。
^ Babity、JM; スター、テレビ; ローズ、AM(1990)。「Caenorhabditiselegansのブリストル株におけるTc1転位とミューテーター活性」。分子遺伝学および一般遺伝学。222(1):65–70。土井:10.1007 / bf00283024。PMID 1978238。S2CID 11275388。 ^ ハリス、LJ; ローズ、AM(1989)。「Caenorhabditiselegansvar。Bristol(株N2)のTc1要素の構造解析」。プラスミド。22(1):10–21。土井:10.1016 / 0147-619X(89)90031-0。PMID 2550981。 ^ Goodier、John L。; デビッドソン、ウィリアムS.(1994)。「Tc1トランスポゾン様配列はサケ科魚類に広く分布している」。分子生物学ジャーナル。241(1):26–34。土井:10.1006 /jmbi.1994.1470。PMID 8051704。 ^ Rosenzweig、B; リャオ、LW; ハーシュ、D(1983)。「C.エレガンス転移因子Tc1の配列」。核酸研究。11(12):4201–4209。土井:10.1093 / nar /11.12.4201。PMC 326035。PMID 6306578。 ^ Schouten、GJ; ヴァンリューネン、HGAM; ベラ、NCV; バレリオ、D。; Plasterk、RHA(1998)。「線虫CaenorhabditiselegansのトランスポゾンTc1がヒト細胞に飛び込む」。核酸研究。26(12):3013–7。土井:10.1093 / nar /26.12.3013。PMC 147650。PMID 9611249。 ^ Vanluenen、H; コロムズ、S; Plasterk、R(1994)。「C.エレガンスにおけるTc3の転位のメカニズム」。セル。79(2):293–301。土井:10.1016 / 0092-8674(94)90198-8。PMID 7954797。S2CID 5804504。 ^ 大隅、T; ベルナップ、WR; ガーリック、B(1995年12月14日)。「人間のマリナートランスポゾン」。自然。378(6558):672 Bibcode:1995Natur.378..672O。土井:10.1038 / 378672a0。PMID 7501013。S2CID 4269186。 ^ ライター、LT; Liehr、T; Rautenstrauss、B; ロバートソン、HM; ルプスキー、JR(1999)。「PRINSによるヒトゲノムにおけるマリナーDNAトランスポゾンの局在化」。ゲノム研究。9(9):839–843。土井:10.1101 /gr.9.9.839。PMC 310809。PMID 10508842。 ^ Jacobson JW、Medhora MM、Hartl DL(1986年11月)。「ショウジョウバエの体細胞的に不安定な転移因子の分子構造」。手順 国立 Acad。科学 アメリカ。83(22):8684–8。Bibcode:1986PNAS … 83.8684J。土井:10.1073 /pnas.83.22.8684。PMC 386995。PMID 3022302。 ^ ロバートソン、HM; マクラウド、EG(1993)。「チチュウカイミバエおよび関連する節足動物を含む、昆虫のマリナー転移因子の5つの主要な亜科」。昆虫分子生物学。2(3):125–39。土井:10.1111 /j.1365-2583.1993.tb00132.x。PMID 9087550。S2CID 11093292。 ^ Lohe AR、Moriyama EN、Lidholm DA、Hartl DL(1995年1月)。「水平感染、垂直不活化、および船員のような転移因子の確率的喪失」。モル。Biol。Evol。12(1):62–72。土井:10.1093 /oxfordjournals.molbev.a040191。PMID 7877497。 ^ Lampe DJ、Witherspoon DJ、Soto-Adames FN、Robertson HM。「セイヨウミツバチ亜科マリナートランスポゾンの4つの異なる順序を表す昆虫系統への最近の水平伝播は、選択が水平伝播中にのみ作用することを示しています」。モル。Biol。Evol。20(4):554–62。土井:10.1093 / molbev / msg069。PMID 12654937。 ^ Mandal PK、カザジアンHH。「SnapShot:脊椎動物のトランスポゾン」。セル。135(1):192–192.e1。土井:10.1016 /j.cell.2008.09.028。PMID 18854165。S2CID 82147。 ^ Carlton JM、Hirt RP、Silva JC、他。。「性感染症の病原体Trichomonasvaginalisのドラフトゲノム配列」。科学。315(5809):207–12。Bibcode:2007Sci … 315..207C。土井:10.1126 /science.1132894。PMC 2080659。PMID 17218520。 ^ Miskey、C。; パップ、B。; メイト、L。; Sinzelle、L。; ケラー、H。; Izsvak、Z。; Ivics、Z。(2007年4月2日)。「古代のマリナーが再び航海する:再構築されたトランスポザーゼによるヒトHsmar1エレメントの転位とトランスポゾン末端でのSETMARタンパク質の活性」。分子生物学および細胞生物学。27(12):4589–4600。土井:10.1128 /MCB.02027-06。PMC 1900042。PMID 17403897。 ^ ギル、エステル; ボッシュ、Assumpcio; ランプ、デビッド; Lizcano、Jose M。; ペラレス、ホセC。; ダノス、オリヴィエ; チロン、ミゲル; コルドー、リチャード(2013年9月11日)。「ヒトマリナートランスポゾンHsmar2の機能的特性」(PDF)。PLOSONE。8(9):e73227。Bibcode:2013PLoSO … 873227G。土井:10.1371 /journal.pone.0073227。PMC 3770610。PMID 24039890。
^ Hartl、D(2001)。「転移因子マリナーの発見」。遺伝学。157(2):471–476。土井:10.1093 /遺伝学/157.2.471。PMC 1461507。PMID 11156971。 ^ ランプ、DJ; Akerley、BJ; ルービン、EJ; Mekalanos、JJ; ロバートソン、HM(1999)。「Himar1マリナートランスポゾンの多活性トランスポザーゼ変異体」。国立科学アカデミーの議事録。96(20):11428–11433。Bibcode:1999PNAS … 9611428L。土井:10.1073 /pnas.96.20.11428。PMC 18050。PMID 10500193。 ^ Gomulski、LM; トルティ、C; Bonizzoni、M; モラリ、D; ライモンディ、E; キャピー、P; ガスペリ、G; マラクリダ、AR。「Ceratitisrosaおよび他のミバエのマリナー要素の新しい基本的な亜科」。分子進化ジャーナル。53(6):597–606。Bibcode:2001JMolE..53..597G。土井:10.1007 / s002390010246。PMID 11677619。S2CID 21289272。 ^ Shao、H; 火、Z。「IS630-Tc1-marinerスーパーファミリーの多様性の拡大:ユニークなDD37Eトランスポゾンの発見とDD37DおよびDD39Dトランスポゾンの再分類」。遺伝学。159(3):1103–15。土井:10.1093 /遺伝学/159.3.1103。PMC 1461862。PMID 11729156。 ^ スミット、AF; リッグス、AD(1996年2月20日)。「ヒトゲノムのティガーとDNAトランスポゾン化石」。アメリカ合衆国科学アカデミー紀要。93(4):1443–8。土井:10.1073 /pnas.93.4.1443。PMC 39958。PMID 8643651。 ^ カソラ、C; ハック、D; Feschotte、C。「分裂酵母および哺乳類におけるセントロメア結合タンパク質へのポゴ様トランスポザーゼの収束的家畜化」。モレキュラーバイオロジーアンドエボリューション。25(1):29–41。土井:10.1093 / molbev / msm221。PMC 2268608。PMID 17940212。 “