Type_II_topoisomerase
II型トポイソメラーゼは、DNAのもつれとスーパーコイルを管理するために、DNAヘリックスの両方の鎖を同時に切断するトポイソメラーゼです。I型トポイソメラーゼとは異なり、ATPの加水分解を使用します。この過程で、これらの酵素は環状DNAの連結数を±2変化させます。
DNAトポイソメラーゼII(ATP加水分解)
他のすべてのIIA型トポイソメラーゼに相同なDNAジャイレースのN末端ATPアーゼおよびトランスデューサードメインの42KDaフラグメントの構造。
識別子
EC番号
5.6.2.2
データベース IntEnz IntEnzビュー
ブレンダ
BRENDAエントリー ExPASy NiceZymeビュー KEGG KEGGエントリー MetaCyc 代謝経路
プリアモス
プロフィール
PDB構造
RCSB PDB PDBe PDBsum索 PMC
記事 PubMed 記事 NCBI タンパク質
コンテンツ
1 関数
2 トポロジーの単純化3 分類 4 構造
4.1 タイプIIA 4.2 タイプIIB
5 作用機序
5.1 ストランド通路 5.2 DNA切断
6 カテネーション
7 阻害
8 バクテリオファージT4ジャイレース
9 参考文献
10 参考文献
11 外部リンク
関数
切断されると、DNAの両端が分離され、2番目のDNA二重鎖が切断部を通過します。継代後、切断されたDNAは再ライゲーションされます。この反応により、II型トポイソメラーゼはDNAループの連結数を2単位増加または減少させ、染色体の解きほぐしを促進します。スーパーコイリングの増加を伴う反応には、2分子のATPが必要です。たとえば、大腸菌や他のほとんどの原核生物で観察されるII型トポイソメラーゼであるDNAジャイレースは、負のスーパーコイルを導入し、連結数を2つ減らします。ジャイレースは細菌の染色体から結び目を取り除くこともできます。ほとんどの原核生物は、ジャイレースに加えて、トポイソメラーゼIVと呼ばれる2番目のIIA型トポイソメラーゼも含んでいます。ジャイレースとトポイソメラーゼIVは、C末端ドメインが異なり、これら2つの酵素の基質特異性と機能性を決定すると考えられています。フットプリントは、140塩基対のフットプリントを形成してDNAを包むジャイレースが負のスーパーコイルを導入するのに対し、28塩基対のフットプリントを形成するトポイソメラーゼIVはDNAを包まないことを示しています。
真核生物のII型トポイソメラーゼはスーパーコイルを導入できません。それは彼らをリラックスさせることしかできません。
IIB型トポイソメラーゼの役割はあまり理解されタイプIIAトポイソメラーゼとは異なり、タイプIIBトポイソメラーゼはDNAトポロジーを単純化できませんが(以下を参照)、タイプIIAトポイソメラーゼといくつかの構造的特徴を共有しています。
トポロジーの単純化
タイプIIAトポイソメラーゼは、複製中の絡み合った娘鎖の分離に不可欠です。この機能は、真核生物ではトポイソメラーゼIIによって、原核生物ではトポイソメラーゼIVによって実行されると考えられています。これらの鎖を分離できないと、細胞死につながります。タイプIIAトポイソメラーゼは、DNAを熱力学的平衡よりも低い状態に緩和する特別な能力を持っています。これは、タイプIA、IB、およびIIBトポイソメラーゼとは異なります。トポロジーの単純化として知られるこの機能は、Rybenkov etalによって最初に特定されました。 ATPの加水分解はこの単純化を促進しますが、この単純化の明確な分子メカニズムはまだ不足しています。この現象を説明するいくつかのモデルが提案されており、IIA型トポイソメラーゼが曲がったDNA二重鎖を認識する能力に依存する2つのモデルが含まれています。生化学、電子顕微鏡、およびDNAに結合したトポイソメラーゼIIの最近の構造は、IIA型トポイソメラーゼがDNAの頂点に結合することを明らかにし、このモデルをサポートしています。
分類
は、新しく見つかったIIBメンバー[doi:10.1093 / nargab / lqz021] この情報を含めるには、セクションを展開して詳細については、トークページをご覧
II型トポイソメラーゼにはIIA型とIIB型の2つのサブクラスが
IIA型トポイソメラーゼには、酵素DNAジャイレース、真核生物のトポイソメラーゼII(topo II)、および細菌のトポイソメラーゼIV(topo IV)が含まれます。これらの酵素は生命のすべての領域にまたがり、機能に不可欠です。
タイプIIBトポイソメラーゼは、構造的および生化学的に異なり、単一のファミリーメンバーであるトポイソメラーゼVI(トポVI)を含みます。タイプIIBトポイソメラーゼは古細菌といくつかの高等植物に見られます。
一部の生物には、トポイソメラーゼIIの2つのアイソフォームがアルファとベータです。癌、トポイソメラーゼII-αは、高度の高い増殖細胞で発現されます。末梢神経鞘腫瘍などの特定の癌では、そのコードされたタンパク質の高発現も患者の生存率の低下に関連しています。
トポイソメラーゼの2つのクラスは、類似した鎖通過メカニズムとドメイン構造を持っていますが(以下を参照)、いくつかの重要な違いもタイプIIAトポイソメラーゼは、4塩基対のオーバーハングを持つ二本鎖切断を形成しますが、タイプIIBトポイソメラーゼは、2塩基対のオーバーハングを持つ二本鎖切断を形成します。さらに、IIA型トポイソメラーゼはDNAトポロジーを単純化することができますが、IIB型トポイソメラーゼはそうではありません。
構造
タイプIIA

の他の例と比較して逆さまに向けられたジャイレースの概略構造。
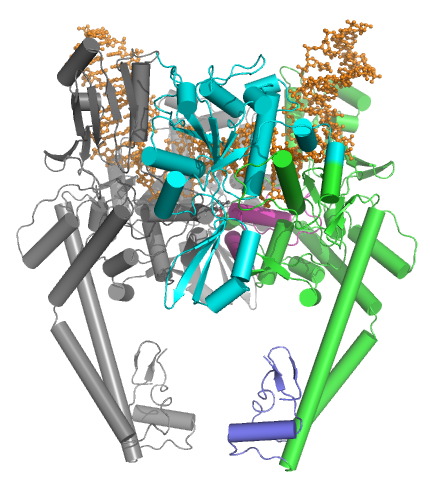
二重にニックが入った34-mer二重鎖DNA(PDB:2RGR)に結合した酵母トポイソメラーゼIIの構造
。Toprimの折り目はシアン色です。DNAはオレンジ色です。HTHはマゼンタ色です。Cゲートは紫色になっています。DNAが不変のイソロイシン(酵母ではIle833)を介して約160度曲がっていることに注意して
タイプIIAトポイソメラーゼは、いくつかの重要なモチーフで構成されています。
N末端GHKL ATPアーゼドメイン(ジャイレース用(HSP)、キナーゼとのMutL)
Toprimドメイン(Rossmann折り畳みの折り両方のII型トポイソメラーゼに存在するサブクラス)、、、IAトポイソメラーゼを入力して、細菌プライマーゼ(DnaGプライマー)
中央のDNA結合コア(構造的にハート型の構造を形成する)、および
可変C末端ドメイン。
真核生物のII型トポイソメラーゼはホモ二量体(A 2)であり、原核生物のII型トポイソメラーゼはヘテロ四量体(A 2 B 2)です。原核生物は1つのポリペプチド(Pfam PF00204)にATPaseドメインとToprimフォールドを持ち、DNA切断コアとCTDは2番目のポリペプチド(Pfam PF00521)にジャイレースの場合、最初のポリペプチドはGyrBと呼ばれ、2番目のポリペプチドはGyrAと呼ばれます。topo IVの場合、最初のポリペプチドはParEと呼ばれ、2番目のポリペプチドはParCと呼ばれます。両方のPfamシグネチャは、一本鎖ユーカヨティックトポイソメラーゼに見られます。
ジャイレースおよび酵母トポイソメラーゼII のN末端ATPaseドメインの構造は、AMPPNP(ATPアナログ)との複合体で解明されており、2つのATPaseドメインが二量体化して閉じたコンフォメーションを形成することを示しています。ジャイレースの場合、構造の中央にかなりの穴があり、Tセグメントを収容すると推定されます。
ATPaseドメインをToprimフォールドにリンクするのは、トランスデューサードメインとして知られるらせん状の要素です。このドメインは、ATPaseドメインのヌクレオチド状態を残りのタンパク質に伝達すると考えられています。このドメインへの変更はトポイソメラーゼ活性に影響を及ぼし、Verdineグループによって行われた構造的研究は、ATP状態がトランスデューサードメインの配向に影響を与えることを示しています。
タンパク質の中心コアには、カタボライト活性化タンパク質のWHDに類似していることが最初に確認されたため、Toprimフォールドと、しばしばCAPドメインと呼ばれるウィングドヘリックスドメイン(WHD)を含むDNA結合コアが含まれています。触媒チロシンはこのWHDにトプリムフォールドはロスマンフォールドであり、DNA切断とDNA再結合に関与するマグネシウムイオンを調整する3つの不変の酸性残基を含みます。酵母トポイソメラーゼIIのトプリムフォールドとDNA結合コアの構造はBergerとWangによって最初に解決され、最初のジャイレースDNA結合コアはMorais Cabral etalによって解決されました。バーガーによって解決された構造は、酵素の機能への重要な洞察を明らかにしました。DNA結合コアは、タワードメインにつながるWHDで構成されています。コイルドコイル領域は、この結晶状態の主要な二量体界面を形成するC末端ドメインにつながります(多くの場合、Cゲートと呼ばれます)。元のトポイソメラーゼII構造は、WHDが遠く離れている状況を示していますが、ジャイレースの構造は、WHDが閉じている閉じたコンフォメーションを示しています。
トポイソメラーゼIIコアは、後にFass etalによるものを含む新しいコンフォメーションで解決されました。そしてDongらによるもの。 Fass構造は、Toprimドメインが柔軟であり、この柔軟性により、ToprimドメインがWHDと協調して、有能な切断複合体を形成できることを示しています。これは最終的にDongらによって実証されました。DNAの存在下で解決された構造。この最後の構造は、トプリムドメインとWHDがIA型トポイソメラーゼと非常によく似た切断複合体を形成し、DNA結合と切断がどのように切り離されるかを示し、構造はDNAが不変のイソロイシン(トポイソメラーゼIIではI833、ジャイレースではI172)。この屈曲のメカニズムは、細菌の2つの構造タンパク質である統合宿主因子(IHF)とHUのメカニズムによく似ています。さらに、DNA結合コアの以前の構造ではCゲートが閉じていましたが、この構造では、2ゲートメカニズムの重要なステップであるゲートが開いています(以下を参照)。
最近では、DNA切断の化学的メカニズムと、抗菌毒によるトポイソメラーゼ阻害の構造的基礎の両方を理解するために、DNA結合構造のいくつかの構造が解明されています。大腸菌DNAジャイレースの最初の完全なアーキテクチャは、原子分解能に近い低温電子顕微鏡法によって解決されました。核タンパク質複合体は、長いDNA二重鎖と、新規の細菌トポイソメラーゼ阻害剤であるゲポチダシンで捕捉されました。
原核生物のトポイソメラーゼのC末端領域は、複数の種で解決されています。ジャイレースのC末端ドメインの最初の構造はCorbettらによって解決されました。そしてトポイソメラーゼIVのC末端ドメインはCorbettらによって解決されました。構造は、核酸をそれ自体の周りに巻き付けることによってDNAを曲げる新しいベータバレルを形成しました。ジャイレースによるDNAの屈曲は、ジャイレースがDNAに負のスーパーコイルを導入する能力の重要なメカニズムとして提案されています。これは、ジャイレースのフットプリントが140塩基対であることを示すフットプリントデータと一致しています。ジャイレースとトポイソメラーゼIVCTDはどちらもDNAを曲げますが、ジャイレースだけが負のスーパーコイルを導入します。
原核生物のトポイソメラーゼのC末端ドメインの機能とは異なり、真核生物のトポイソメラーゼIIのC末端領域の機能はまだ明らかではありません。研究は、この領域がリン酸化によって調節され、これがトポイソメラーゼ活性を調節することを示唆していますが、これを調査するためにさらに研究を行う必要が
タイプIIB

酵母の例と同様の向きのトポVI(PDB:2Q2E)の構造
。チェーンの色は異なります。Toprimドメインは上にあり、ATPaseドメインは下にそれぞれがDNAゲートを形成します。
タイプIIBトポイソメラーゼの構成は、すべてのタイプIIBが2つの遺伝子を持ち、ヘテロ四量体を形成することを除いて、タイプIIAの構成と似ています。topo VI-B(gyrBに似ているため)と呼ばれる1つの遺伝子には、ATPaseドメイン、トランスデューサードメイン(Pfam PF09239)、およびC末端Igフォールド様H2THドメイン(Pfam PF18000)が含まれています。topo VI-A(Pfam PF04406)と呼ばれる2番目の遺伝子には、WHDとToprimドメインが含まれています。
topo VIBのATPaseドメインは複数のヌクレオチド状態で解決されました。これは、topo IIおよびMutLのGHKLドメインのドメインと非常によく似ており、ヌクレオチドの状態(ADP対ATP)がトランスデューサードメイン(および1MX0)の方向に影響を与えることを示しています。
トポVI-Aの構造はBergeratらによって解決されました。 HTHとToprimの折り畳みがトポIIAのそれと比較して新しいコンフォメーションを持っていたことを示しています。
トポVIA / B複合体の最近の構造が解明され、2ゲートメカニズムで予測される2つの状態であるオープンコンフォメーションとクローズドコンフォメーションが示されました(以下を参照)。これらの構造は、1つはX線結晶構造で、もう1つは小角X線散乱(SAXS)再構成であり、ATPaseドメインが開いているか閉じているかを示しています。
作用機序
ストランド通路
IIA型トポイソメラーゼは、「2ゲート」メカニズム(これは歴史的な表記法ですが)、生化学および構造的研究によってサポートされるメカニズムを介して動作します。
ゲートまたはGセグメントと呼ばれるDNAの鎖は、中央のDNA結合ゲート(DNAゲート)によって結合されます。トランスポートまたはTセグメントと呼ばれるDNAの2番目の鎖は、ATPの2つの分子が結合するときにN末端ATPaseドメイン(ATPaseゲート)の二量体化によって捕捉されます。ATPの加水分解と無機リン酸塩の放出は、触媒チロシンがDNAの5 ‘末端と共有結合ホスホチロシン結合を形成するため、Gセグメントの切断につながります。これにより、Gセグメントに4ベースのオーバーハングと2本鎖の切断が作成されます。DNA結合ゲートが分離すると、TセグメントはGセグメントを介して転送されます。Gセグメントは密閉されており、C末端ゲート(またはCゲート)が開き、Tセグメントの解放が可能になります。製品ADPをリリースすると、システムがリセットされ、2番目のTセグメントをキャプチャできるようになります。
タイプIIBトポイソメラーゼは、タンパク質がGセグメントで2塩基のオーバーハングを形成し、C末端ゲートが完全に欠落していることを除いて、同様の方法で動作します。
DNA切断
鎖通過メカニズムでは、DNAの切断がTセグメントがGセグメントを介して移動できるようにするための鍵となります。IIA型トポイソメラーゼによるDNA切断のメカニズムは、最近、多くの生化学的および構造生物学研究の焦点となっています。
カテネーション
カテネーションは、2本の環状DNA鎖が鎖状のリンクのように一緒にリンクされるプロセスです。これは、DNA複製の後に発生します。この場合、2つの一本鎖が連結され、複製はできますが、2つの娘細胞に分離することはできません。タイプIIトポイソメラーゼは二本鎖を切断するため、この状態を修正でき(タイプIトポイソメラーゼは、すでに一本鎖ニックが存在する場合にのみこれを行うことができます)、正しい染色体番号を娘細胞に残すことができます。真核生物の線形DNAは非常に長いため、終わりがないと考えることができます。同じ理由でII型トポイソメラーゼが必要です。
阻害
トポイソメラーゼ阻害剤
II型トポイソメラーゼを標的とする小分子は、阻害剤と毒物の2つのクラスに分類されます。増殖中の真核細胞に頻繁に存在するため、II型トポイソメラーゼの阻害剤は広く研究され、抗がん剤として使用されてきました。
II型トポイソメラーゼの阻害剤には、HU-331、ICRF-187、ICRF-193、およびミチンドマイドが含まれます。これらの分子は、ATPの非競合的阻害剤として作用することによりATPase活性を阻害することによって機能します。これは、リンズリーグループによって実施された構造研究および生化学的研究を通じて示されています。
II型トポイソメラーゼの毒物には、ドキソルビシン、エトポシド、ノボビオシン、キノロン(シプロフロキサシンを含む)、およびテニポシドが含まれます。これらの小分子は、DNA-タンパク質複合体を標的としています。これらの分子のいくつかは切断の増加をもたらしますが、エトポシドなどの他の分子は再結合を阻害します。
実験的抗腫瘍薬m-AMSA(4 ‘-(9’-アクリジニルアミノ)メタンスルホン-m-アニシジド)も2型トポイソメラーゼを阻害します。
トポイソメラーゼ毒は、抗癌療法と抗菌療法の両方として広く使用されています。シプロフロキサシンなどの抗菌性化合物は細菌ジャイレースを標的としますが、真核生物のIIA型トポイソメラーゼを阻害することはできません。さらに、薬剤耐性菌はしばしばジャイレース(大腸菌ではSerine79Alanine)に点突然変異を持っており、キノロンを無効にします。最近の構造研究により、この残留物に依存しなくなったため、薬剤耐性菌に対して有効な化合物が発見されました。
バクテリオファージT4ジャイレース
バクテリオファージ(ファージ)T4ジャイレース(II型トポイスメラーゼ)は、遺伝子39、52、およびおそらく60の産物からなるマルチサブユニットタンパク質です。 これは、負または正の超らせんDNAの弛緩を触媒し、ファージに使用されます。E.coliバクテリオファージ宿主の感染中のDNA複製。ファージ遺伝子52タンパク質は大腸菌ジャイレースgyrAサブユニットと相同性を共有し、ファージ遺伝子39タンパク質はgyrBサブユニットと相同性を共有します。宿主の大腸菌DNAジャイレースはファージT4遺伝子産物の喪失を部分的に補うことができるため、遺伝子39、52、または60のいずれかに欠陥のある変異体は、ファージDNA複製を完全に無効にするのではなく、その開始を遅らせる。 DNAの伸長速度は、そのような突然変異体感染症では野生型よりも遅くはない。遺伝子39、52または60に欠陥のある突然変異体は、遺伝子組換えの増加、ならびに塩基置換および欠失突然変異の増加を示し、宿主補償DNA合成が野生型ファージによって指示されるものよりも精度が低いことを示唆している。遺伝子39に欠陥のある変異体は、ファージ染色体の複数のコピーが存在する場合、DNA複製の開始後のファージ感染の段階で紫外線照射による不活化に対する感受性の増加を示します。遺伝子39、52、および60に欠陥のある変異体は、さまざまなタイプのDNA損傷に対処できる組換え修復の一形態である多重度再活性化を実行する能力が低下しています。感染していない大腸菌のゲノムによって特定されたジャイレースも、RecAタンパク質によって駆動される相互鎖交換の開始点を提供することによって組換え修復に関与しているようです。
参考文献
^ Rybenkov VV、Ullsperger C、Vologodskii AV、Cozzarelli NR(1997年8月)。「II型トポイソメラーゼによる平衡値以下のDNAトポロジーの単純化」。科学。ニューヨーク州ニューヨーク277(5326):690–3。土井:10.1126 /science.277.5326.690。PMID 9235892。
^ Vologodskii AV、Zhang W、Rybenkov VV、Podtelezhnikov AA、Subramanian D、Griffith JD、Cozzarelli NR。「II型DNAトポイソメラーゼによるトポロジー単純化のメカニズム」。アメリカ合衆国科学アカデミー紀要。98(6):3045–9。Bibcode:2001PNAS … 98.3045V。土井:10.1073 /pnas.061029098。PMC 30604。PMID 11248029。 ^ Reece RJ、Maxwell A(1991年1月)。「DNAジャイレース:構造と機能」。生化学および分子生物学における批評的レビュー。26(3–4):335–75。土井:10.3109 / 10409239109114072。PMID 1657531。 ^ Buhler C、Lebbink JH、Bocs C、Ladenstein R、Forterre P。「DNAトポイソメラーゼVIは、2ヌクレオチドのオーバーハングを伴うATP依存性の二本鎖切断を生成します」。Journal of BiologicalChemistry。276(40):37215–22。土井:10.1074 /jbc.M101823200。PMID 11485995。S2CID 24354635。 ^ PDB:1zvt ;
Corbett KD、Schoeffler AJ、Thomsen ND、Berger JM。「DNAトポイソメラーゼIVにおける基質特異性の構造的基礎」。分子生物学ジャーナル。351(3):545–61。土井:10.1016 /j.jmb.2005.06.029。PMID 16023670。 ^ Wigley DB、Davies GJ、Dodson EJ、Maxwell A、Dodson G(1991年6月)。「DNAジャイレースBタンパク質のN末端フラグメントの結晶構造」。ネイチャー。351(6328):624–9。Bibcode:1991Natur.351..624W。土井:10.1038 / 351624a0。PMID 1646964。S2CID 4373125。 ^ PDB:1PVG ;
Classen S、Olland S、Berger JM。「トポイソメラーゼIIATPase領域の構造と化学療法剤ICRF-187による阻害のメカニズム」。アメリカ合衆国科学アカデミー紀要。100(19):10629–34。Bibcode:2003PNAS..10010629C。土井:10.1073 /pnas.1832879100。PMC 196855。PMID 12963818。 ^ Wei H、Ruthenburg AJ、Bechis SK、Verdine GL。「ヒトIIA型DNAトポイソメラーゼのATPaseドメインにおけるヌクレオチド依存性ドメインの動き」。Journal of BiologicalChemistry。280(44):37041–7。土井:10.1074 /jbc.M506520200。PMID 16100112。S2CID 35186716。 ^ Aravind L、Leipe DD、Koonin EV(1998年9月)。「トプリム-IA型およびII型トポイソメラーゼ、DnaG型プライマーゼ、OLDファミリーヌクレアーゼおよびRecRタンパク質の保存された触媒ドメイン」。核酸研究。26(18):4205–13。土井:10.1093 / nar /26.18.4205。PMC 147817。PMID 9722641。 ^ PDB:1BGW ;
バーガーJM、ガンブリンSJ、ハリソンSC、ワンJC(1996年1月)。「DNAトポイソメラーゼIIの構造とメカニズム」。ネイチャー。379(6562):225–32。Bibcode:1996Natur.379..225B。土井:10.1038 / 379225a0。PMID 8538787。S2CID 4360011。 ^ PDB:1AB4 ;
Morais Cabral JH、Jackson AP、Smith CV、Shikotra N、Maxwell A、Liddington RC(1997年8月)。「DNAジャイレースの破壊-再結合ドメインの結晶構造」。ネイチャー。388(6645):903–6。Bibcode:1997Natur.388..903M。土井:10.1038 / 42294。PMID 9278055。S2CID 4320715。 ^ PDB:1BJT ;
Fass D、Bogden CE、Berger JM(1999年4月)。「トポイソメラーゼIIの四次変化は、2本のDNA鎖の直交運動を指示する可能性があります」。自然構造生物学。6(4):322–6。土井:10.1038 / 7556。PMID 10201398。S2CID 947461。 ^ PDB:2RGR ;
ドンKC、バーガーJM。「IIA型トポイソメラーゼによるゲートDNA認識と屈曲の構造的基盤」。ネイチャー。450(7173):1201–5。Bibcode:2007Natur.450.1201D。土井:10.1038 / nature06396。PMID 18097402。S2CID 1756317。 ^ Vanden Broeck A、Lotz C、Ortiz J、Lamour V。「完全な大腸菌DNAジャイレース核タンパク質複合体の低温電子顕微鏡構造」。ネイチャーコミュニケーションズ。10(1):4935. Bibcode:2019NatCo..10.4935V。土井:10.1038 / s41467-019-12914-y。PMC 6821735。PMID 31666516。 ^ PDB:1SUU ;
Corbett KD、Shultzaberger RK、Berger JM。「DNAジャイレースAのC末端ドメインはDNAを曲げるベータ風車の折り目を採用しています」。アメリカ合衆国科学アカデミー紀要。101(19):7293–8。Bibcode:2004PNAS..101.7293C。土井:10.1073 /pnas.0401595101。PMC 409912。PMID 15123801。 ^ PDB:1MU5 ;
Corbett KD、Berger JM。「トポイソメラーゼVI-Bサブユニットの構造:II型トポイソメラーゼのメカニズムと進化への影響」。EMBOジャーナル。22(1):151–63。土井:10.1093 / emboj / cdg008。PMC 140052。PMID 12505993。 ^ Bergerat A、de Massy B、Gadelle D、Varoutas PC、Nicolas A、Forterre P(1997年3月)。「古細菌由来の非定型トポイソメラーゼIIと減数分裂組換えへの影響」。ネイチャー。386(6623):414–7。Bibcode:1997Natur.386..414B。土井:10.1038 / 386414a0。PMID 9121560。S2CID 4327493。 ^ PDB:2Q2E ;
Corbett KD、Benedetti P、Berger JM。「トポイソメラーゼVIのホロ酵素集合とATPを介したコンフォメーションダイナミクス」。自然構造および分子生物学。14(7):611–9。土井:10.1038 / nsmb1264。PMID 17603498。S2CID 2159631。 ^ Roca J、Wang JC(1994年5月)。「II型DNAトポイソメラーゼによるDNA輸送:2ゲートメカニズムを支持する証拠」。セル。77(4):609–16。土井:10.1016 / 0092-8674(94)90222-4。PMID 8187179。S2CID 19776252。 ^ バーガーJM、ワンJC(1996年2月)。「DNAトポイソメラーゼIIの構造とメカニズムの最近の開発」。構造生物学における現在の意見。6(1):84–90。土井:10.1016 / s0959-440x(96)80099-6。PMID 8696977。 ^ アルバーツB(2014-11-18)。細胞の分子生物学(第6版)。ニューヨーク州ニューヨーク。ISBN 978-0-8153-4432-2。OCLC 887605755。
^ Willmore E、de Caux S、Sunter NJ、Tilby MJ、Jackson GH、Austin CA、Durkacz BW。「新規のDNA依存性プロテインキナーゼ阻害剤であるNU7026は、白血病の治療に使用されるトポイソメラーゼII毒の細胞毒性を増強します」。血。103(12):4659–65。土井:10.1182 / blood-2003-07-2527。PMID 15010369。 ^ Liu LF、Liu CC、Alberts BM(1979年10月)。「T4DNAトポイソメラーゼ:T4バクテリオファージDNA複製の開始に不可欠な新しいATP依存性酵素」。ネイチャー。281(5731):456–61。Bibcode:1979Natur.281..456L。土井:10.1038 / 281456a0。PMID 226889。S2CID 4343962。 ^ Stetler GL、King GJ、Huang WM(1979年8月)。「特定のDNA複製に必要なT4DNA遅延タンパク質は、ATP依存性のDNAトポイソメラーゼ活性を持つ複合体を形成します」。アメリカ合衆国科学アカデミー紀要。76(8):3737–41。Bibcode:1979PNAS … 76.3737S。土井:10.1073 /pnas.76.8.3737。PMC 383908。PMID 226976。 ^ マッカーシーD(1979年1月)。「バクテリオファージT4DNA複製のジャイレース依存性開始:大腸菌ジャイレースとノボビオシン、クメルマイシンおよびファージDNA遅延遺伝子産物との相互作用」。分子生物学ジャーナル。127(3):265–83。土井:10.1016 / 0022-2836(79)90329-2。PMID 372540。 ^ 黄WM(1986年9月)。「T4DNAトポイソメラーゼの52タンパク質サブユニットは、ジャイレースのgyrAタンパク質と相同です」。核酸研究。14(18):7379–90。PMC 311757。PMID 3020513。 ^ 黄WM(1986年10月)。「II型DNAトポイソメラーゼ遺伝子のヌクレオチド配列。バクテリオファージT4遺伝子39」。核酸研究。14(19):7751–65。土井:10.1093 / nar /14.19.7751。PMC 311794。PMID 3022233。 ^ McCarthy D、Minner C、Bernstein H、Bernstein C(1976年10月)。「野生型ファージT4とDNA遅延琥珀変異体のDNA伸長率と成長点分布」。分子生物学ジャーナル。106(4):963–81。土井:10.1016 / 0022-2836(76)90346-6。PMID 789903。 ^ Mufti S、Bernstein H(1974年10月)。「バクテリオファージT4のDNA遅延変異体」。ウイルス学ジャーナル。14(4):860–71。土井:10.1128 /JVI.14.4.860-871.1974。PMC 355592。PMID 4609406。 ^ ハイマンP(1993年8月)。「バクテリオファージT4におけるLuria-Latarjet効果の遺伝学:複数のDNA修復経路の関与の証拠」。遺伝子研究。62(1):1–9。土井:10.1017 / s0016672300031499。PMID 8405988。 ^ Miskimins R、Schneider S、Johns V、Bernstein H(1982年6月)。「ファージT4の多様性再活性化におけるトポイソメラーゼの関与」。遺伝学。101(2):157–77。doi:10.1093 / genetics / 101.2.157(2021年10月31日非アクティブ)。PMC 1201854。PMID 6293912。 ^ Cassuto E(1984年9月)。「ジャイレースとRecAタンパク質の複合作用による共有結合で閉じたヘテロ二本鎖DNAの形成」。EMBOジャーナル。3(9):2159–64。土井:10.1002 /j.1460-2075.1984.tb02106.x。PMC 557658。PMID 6092061。
参考文献
王JC。「DNAトポイソメラーゼの細胞の役割:分子の展望」。ネイチャーレビュー。分子細胞生物学。3(6):430–40。土井:10.1038 / nrm831。PMID 12042765。S2CID 205496065。
外部リンク
コモンズには、II型DNAトポイソメラーゼに関連するメディアが
DNA +トポイソメラーゼ、+ Type + II、米国国立医学図書館のMedical Subject Headings(MeSH)
![]()
生物学ポータル”