Tyrannosaurinae
tyrannosaurines(ティラノサウルス亜科は)の絶滅サブファミリーであるティラノサウルス科ファミリーのcoelurosaurian 獣脚類の 恐竜少なくとも三つの部族と複数から成る属。これらの属のすべての化石は、北アメリカ西部と東アジアの白亜紀後期の堆積物で発見されています。関連する亜科アルバートサウルス亜科と比較すると、アリオラミンは比較すると繊細でしたが、チラノサウルスは全体的に頑強で大きくなっています。この亜科には、最も古い既知のティラノサウルス属Lythronaxも含まれますティラノサウルスグループの最年少で最後に知られているメンバーの1人です。
チラノサウリン
時間範囲:
白亜紀後期、
80.6〜66 MaPreꞒ O K
Pg アリオラムスの頭蓋骨。
ティラノサウルスの頭蓋骨。
科学的分類
王国: 動物界
門: 脊索動物
クレード:
恐竜
クレード:
竜盤類
クレード:
獣脚類
クレード:
†パンティラノサウリア
クレード: † Eutyrannosauria 家族: †ティラノサウルス科
亜科: †ティラノサウルスオズボーン、1906年
タイプ種
†ティラノサウルスレックス
オズボーン、1905年
サブグループ
†ビスタヒエヴェルソル?
†ダイナモテラー
†リトロナクス
†ナヌークサウルス
†テラトフォネウス
†アリオラミニ
†アリオラムス
†キアンゾウサウルス † Daspletosaurini †ダスプレトサウルス
†タナトテリステス
†ティラノサウルス
†タルボサウルス
†ティラノサウルス
†ズケンティラヌス
コンテンツ
1 発見の歴史
2 自然史
2.1 説明 2.2 分布
3 分類学
4 参考文献
発見の歴史

デイノドンの歯、最も初期の既知のティラノサウルスが残っています
ティラノサウルスの最初の残骸は、カナダ地質調査部が主導する遠征中に発見されました。この調査では、多数の散在する歯が見つかりました。これらの特徴的な恐竜の歯は、1856年にジョセフ・ライディによってデイノドン(「ひどい歯」)という名前が付けられました。1892年、エドワード・ドリンカー・コープは、孤立した脊椎の形でより多くのティラノサウルスの材料を説明し、この動物にManospondylusgigasという名前を付けました。この発見は1世紀以上にわたってほとんど見過ごされていましたが、2000年代初頭に、この資料が実際にティラノサウルスレックスに属し、名前が優先されていることが発見されたときに論争を引き起こしました。 1905年の後半、ヘンリーフェアフィールドオズボーンは、バーナムブラウンが率いる、1902年のアメリカ自然史博物館の遠征中にモンタナとワイオミングで収集された2つのティラノサウルス標本について説明しました。当初、オズボーンはこれらを別個の種と見なしていました。最初に、彼はDynamosaurus imperiosus(「皇帝の力のトカゲ」)と名付け、2番目の名前はTyrannosaurus rex(「王の暴君のトカゲ」)と名付けました。1年後、オズボーンはこれら2つの標本が実際には同じ種からのものであることを認識しました。ダイナモサウルスが最初に発見されたという事実にもかかわらず、ティラノサウルスという名前は、両方の標本を説明する彼の元の記事の1ページ前に登場していました。したがって、国際動物命名規約(ICZN)によると、ティラノサウルスという名前が使用されました。
第二はtyrannosaurinesの代表的な説明、タルボサウルスは、(本来のアジア代表として説明ティラノサウルス大頭蓋骨を関節中に回収された後の)1955年にあったソ連に-Mongolian遠征ゴビ砂漠モンゴルにおけるウムヌゴビ県1946年にホロタイプでした名前のティラノサウルスバターでエビニー・マリーバとしてティラノサウルスバター。タルボサウルス属は、1948年と1949年にタルボサウルスエフレモビと同じ遠征で発見された頭蓋骨と骨格の残骸を含む標本であるPIN 551–2に基づいて同じ年に記述されました。 Tyが登場したのは1965年のことでした。バタールとタ。efremoviは、同じ種であった後者は、若い動物である、とは異なるティラノサウルスはによって認識AKロジェストヴェンスキーのような種を再結合タルボサウルスのバター。
1970年代に2つの属の記述を見ました。1970年にDaspletosaurusが出版され、ホロタイプCMN 8506は、頭蓋骨、肩、前肢、骨盤、大腿骨、首、胴体、腰からのすべての椎骨を含む部分的な骨格で構成されています。最初の11の尾の椎骨。それは、近くの1921年に発見されたSteveville、アルバータにより、チャールズ・モートラム・スタンバーグ、それはの新種だと思った、ゴルゴサウルス、しかしによって新属であることが判明したデール・ラッセル前述のCMN 8506.使用して、 、第二のアリオラムス、 1976年にセルゲイクルザーノフによって記述されたホロタイプ(PIN 3141/1)は、1970年代初頭にソビエトとモンゴルのゴルゴサウルスへの合同遠征で発見された3つの中足骨に関連する部分的な頭蓋骨であり、これらはノゴンツァフとして知られる地域に残っています。モンゴルのバヤンコンゴール州、ネメグト層。
1977年から2009年にかけて、いくつかの属の出版物が見られました。アジアでは、彼らが含まShanshanosaurus(1977)、 Maleevosaurus(1992)、とラプトレックス(2009)、北米で見ながらナノティラヌス(1988)、 DinotyrannusとStygivenator(1995)。しかしながら、これらの動物の残骸は未成熟または幼い個体であるため、これらの属は物議を醸しています。2021年の時点で、アジアの標本はタルボサウルスの初期成長段階であり 、北米の標本はティラノサウルスの標本であることが広く理解されています。

リトロナクス(A)と
テラトフォネウス標本(B)のホロタイプの残骸を
示す骨格図
。N–Pは前者の選択された骨を示します
有効な属は2010年代まで名前が付けられませんでした。そこでは、2011年にトーマスD.カーとその同僚によるテラトフォネウスの出版が発表されました。化石は、ユタ州南部のカイパロウィッツ層で最初に発見されました。その後、同じ層の化石が発見され、属として特定されました。アルゴン-アルゴン放射年代測定は、カイパロウィッツ層が76.1〜74.0百万年前、白亜紀後期のカンパニアン期に堆積したことを示しています。この日付は、テラトフォネウスが白亜紀後期のカンパニアン期の真ん中に住んでいたことを意味します。同じ年、Zhuchengtyrannusは、諸城恐竜博物館に収容されているほぼ完全な右上顎骨および関連する左歯(下顎、両方とも歯)であるホロタイプZCDMV0031に基づいてDavidWEHoneらによって命名されました。 2014年には、アメリカ合衆国アラスカのノーススロープのプリンスクリーク層で見つかった最北端のティラノサウルスであるナヌークサウルスがやってきた。 同じ年に、下顎がすべての歯を失った(化石化中に失われた)ほぼ完全な頭蓋骨、9つの頸椎、3つの背椎、18の尾骨からなる部分的な亜成人から知られるキアンゾウサウルスも発表した。椎骨、両方scapulocoracoids、部分的腸骨、および損なう左後肢大腿骨、脛骨、腓骨、レンゲと踵骨および中足骨III及びIVを。リトロナクス、ティラノサウルス科の最古の既知のメンバーは、2009年に発見されたほぼ完全な試料からのMark A.ローウェンらによって2013年に記載されたWahweap形成の大階段エスカランテ国定。
自然史
説明

人間と比較した
A.remotusのサイズ
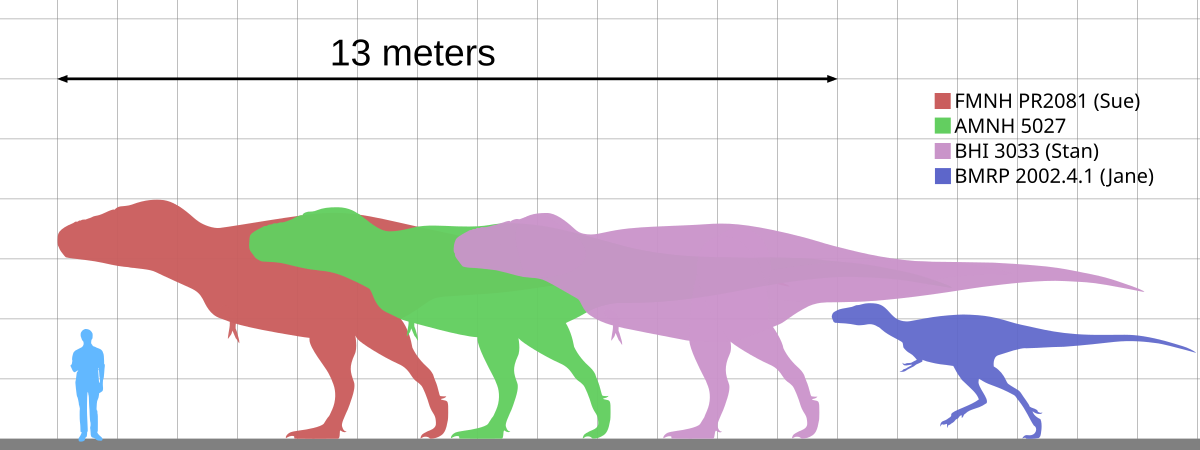
標本「スー」、AMNH 5027、「スタン」、および「ジェーン」は、人間に合わせて拡大縮小されます。
アルバートサウルスと比較して、チラノサウルスはより重く構築され、より大きくなりました。alioraminの属Qianzhousaurusとアリオラムスは、彼らがalbertosaurinesに構築され、longirostrine snoutsを持っているにより同等であったとして、しかし、例外でした。アルバートサウルスと同様に、チラノサウルスも異歯性の歯列、獲物を捕まえて殺すための大きな頭のデザイン、短いジダクチルの腕を持っていた。ティラノサウルス(およびおそらくタルボサウルス)の成長段階に基づいて、ティラノサウルスは個体発生的に変化し、繊細または細く、セミロンギロストリンの未成熟から丈夫で頭の重い成虫になりました。これは、これらの動物が成長するにつれて、異なる生態学的ニッチを占めることを意味します。初期のティラノサウルスが羽毛を持っているという化石の証拠があるが、 ティラノサウルスにおけるそのような構造の証拠は、2017年のベルらの研究がティラノサウルスの羽毛のある外皮の支持を発見しなかったため、物議を醸している。この研究では、頭蓋骨後部の体のさまざまな領域に、ワニに似たパターンで広く分散していることがわかった、小さい皮膚の印象を使用しました。また、の鼻研究することによって2017年ワニ類推トーマス・カーや同僚ダスプレトサウルスは、彼らが皮膚の下に、感覚、感覚ニューロンピットを持つ大規模なスケールを持って提案この概念は、複数の歯のリップカバーを示唆した他の著者から挑戦されています。
分布
チラノサウリンの時間的範囲は、白亜紀後期のカンパニアン期のほぼ8,060万年前から、マーストリヒチアン期の6,600万年前まででした。化石は、現在の東アジアと北アメリカ西部のさまざまな地層で発見されています。アジアのアリオラミンはティラノサウルスの最も基本的なグループですが、北アメリカで見つかったアルバートサウルスや他のユーティラノサウルスのティラノサウルスの地理的配置は、ティラノサウルスが北米起源であることを大いに示唆しています。
分類学
2010年代に先立ち、tyrannosaurinesの関係は最高のものとして理解されたティラノサウルスはの姉妹分類群であることタルボサウルス。順番にこれらの2つの属は姉妹分類群だったダスプレトサウルスによるフォローアリオラムス。対立仮説があったフィル・カリー提案とその同僚(2003)ダスプレトサウルスをより密接に関連するとタルボサウルスとアリオラムスに比べティラノサウルス頭蓋の特徴に基づきます。しかしながら、この関係は最近の研究では発見され 1988年、グレゴリー・ポールは、アリオラムスを除く当時のすべてのティラノサウルスをティラノサウルスの種と見なしました。の第二版では恐竜にプリンストンフィールドガイド2016年に出版され、ポールは、以下を含むだけでなく、このような考えを続けるBistahieversor、Teratophoneus、リトロナクス、およびNanuqsaurusを同様属に。しかしながら、この複数種のティラノサウルス分類は、ほとんどの古生物学者によって広く受け入れられいくつかの系統発生研究では、ビスタヒエヴェルソルはティラノサウルス科に入れ子になっていますが 、代わりにティラノサウルス科の姉妹分類群として回収されることがほとんどです。
2021年の時点で、チラノサウリンには少なくとも3つの系統が基礎最もクレードは部族でAlioramini、発散する第二のクレードは、米国南西の分類群を含む名前クレードあるDynamoterror、リトロナクス、及びTeratophoneusを。彼らは、ナヌークサウルスとダスプレトサウリーニ族とティラノサウリーニ族からなる分岐群の姉妹です。
以下のクラドグラムは、Vorisらによって実行された系統発生分析の結果です。(2020):Eutyrannosauria
ドリプトサウルスaquilunguis
アパラチオサウルスモンゴメリエンシス
ビスタヒエヴェルソルsealeyi
ティラノサウルス科
アルバートサウルス科 ゴルゴサウルスlibratus
アルバートサウルス肉食動物
ティラノサウルス科
アリオラミニ キアンゾウサウルスsinensis
lioramus remotus
アリオラムスアルタイ テラトフォネウスカリー
Dynamoterrorダイナスト
リトロナクスが主張する
ナヌークサウルスhoglundi Daspletosaurini Thanatotheristes degrootorum
ダスプレトサウルストロサス
ダスプレトサウルスホルネリ
ティラノサウルス ズケンティラヌスマグナス
タルボサウルスbataar
ティラノサウルス・レックス
参考文献
^ Holtz、Thomas R. Jr.(2012) Dinosaurs:The Most Complete、Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages、 Winter2011付録。 ウェイバックマシンで2017年8月12日にアーカイブ ^ Breithaupt、BH; サウスウェル、EH; マシューズ、NA(2005年10月18日)。「ティラノサウルスレックスの100年を祝って:Manospondylus gigas、Ornithomimus grandis、およびDynamosaurus imperiosus、西部でのティラノサウルスレックスの最も初期の発見」。プログラムによる要約。2005ソルトレイクシティ年次総会。37。アメリカ地質学会。NS。406. 2019年10月22日にオリジナルからアーカイブされました。
^ Breithaup、BH; サウスウェルEH; マシューズNA(2006)。「ダイナモサウルスインペリオサスとワイオミング州と西部でのティラノサウルスレックスの初期の発見」。ニューメキシコ自然史科学博物館紀要。35:257–258。
^ マレエフ、エフゲニーA.(1955)。「モンゴルの巨大な肉食恐竜」。Doklady Akademii NaukSSSR。104(4):634–637。
^ マレエフ、EA(1955)。FJアルコックによる翻訳。「モンゴルの白亜紀後期からの新しい肉食恐竜」(PDF)。Doklady Akademii NaukSSSR。104(5):779–783。
^ Rozhdestvensky、Anatoly K.(1965)。「アジアの恐竜の成長の変化とそれらの分類法のいくつかの問題」。古生物学ジャーナル。3:95〜109。
^ ラッセル、デールA.(1970)。「西部カナダの後期白亜紀からのティラノサウルス」。古生物学の国立自然科学博物館の出版物。1:1〜34。
^ Kurzanov、SergeiM。「モンゴルのNogon-Tsavの後期白亜紀からの新しいカルノサウルス」。ソビエトとモンゴルの合同古生物学遠征取引(ロシア語)。3:93〜104。
^ 董枝明(1977)。「中国新疆ウイグル自治区トルファンの恐竜遺跡について」。Vertebrata PalAsiatica(中国語)。15:59–66。
^ カーペンター、ケン。(1992)。「アジアと北アメリカのティラノサウルス(恐竜)」。Mateerでは、Niall J。; ペイジ、チェン(編)。非海洋白亜紀の地質学の側面。北京:チャイナオーシャンプレス。pp。250–268。
^ セレノ、P。; タン、L。; ブルサット、SL; Kriegstein、HJ; 趙、X。; クローワード、K。(2009)。「ティラノサウルスの骨格デザインは、最初は小さな体のサイズで進化しました」。科学。326(5951):418–422。Bibcode:2009Sci … 326..418S。土井:10.1126 /science.1177428。PMID 19762599。S2CID 1953405。 ^ バッカー、RT; ウィリアムズ、M。; カリー、PJ(1988)。「モンタナの最新の白亜紀からのピグミーティラノサウルスの新属、ナノティラヌス」。ハンタリア。1:1〜30。
^ オルシェフスキー、G。(1995)。「ティラノサウルスの起源と進化」。極限最前線【ディノ最前線】。9–10:92–119。
^ Carr、Thomas D.(1999)。「ティラノサウルス科(恐竜、コエルロサウルス)の頭蓋顔面個体発生」。Journal of VertebratePaleontology。19(3):497–520。土井:10.1080 /02724634.1999.10011161。
^ カリー、フィリップJ。; 董枝明(2001)。「中国の白亜紀後期からの幼若ティラノサウルス(獣脚類、恐竜)、シャンシャノサウルスhuoyanshanensisに関する新しい情報」。地球科学のカナダジャーナル。38(12):1729–1737。Bibcode:2001CaJES..38.1729C。土井:10.1139 / cjes-38-12-1729。
^ ファウラー、DW; ウッドワード、HN; フリードマン、EA; ラーソン、PL; ホーナー、JR(2011)。「「Raptorexkriegsteini」の再分析:モンゴルからの幼いティラノサウルス恐竜」。PLOSONE。6(6):e21376。Bibcode:2011PLoSO … 6E1376F。土井:10.1371 /journal.pone.0021376。PMC 3126816。PMID 21738646。 ^ ウッドワード、ホリーN。; トレメイン、ケイティ; ウィリアムズ、スコットA。; ザンノ、リンゼイE。; ホーナー、ジョンR。; ネイサン・ミルボルド(2020)「ティラノサウルスレックスの成長:骨組織学はピグミー「ナノティラヌス」に反論し、幼若ティラノサウルスの個体発生ニッチ分割をサポートします」。サイエンスアドバンシス。6(1):eaax6250。Bibcode:2020SciA …. 6.6250W。土井:10.1126 /sciadv.aax6250。ISSN 2375年から2548年。PMC 6938697。PMID 31911944。 ^ Carr、TD(2020)。「複数の証拠から得られたティラノサウルスレックスの高解像度成長シリーズ」。PeerJ。8:e9192。土井:10.7717 /peerj.9192。
^ Carr、TD; ウィリアムソン、TE; ブリット、BB; シュタットマン、K。(2011)。「アメリカ南西部の白亜紀後期(カンパニアン後期)における高い分類学的および形態学的なティラノサウルスの多様性と、ユタ州のカイパロウィッツ層からの新しい短い頭蓋骨のティラノサウルスの証拠」。Naturwissenschaften。98(3):241–246。Bibcode:2011NW ….. 98..241C。土井:10.1007 / s00114-011-0762-7。PMID 21253683。S2CID 13261338。 ^ 研ぎ澄まされた、DWE; 王、K。; サリバン、C。; 趙、X。; Chen、S。; Li、D。; Ji、S。; ジ、Q。; Xu、X。(2011)。「中国の白亜紀後期からの新しい、大きなティラノサウルス獣脚類」。白亜紀の研究。32(4):495–503。土井:10.1016 /j.cretres.2011.03.005。
^ フィオリロ、AR ; Tykoski、RS(2014)。ドッドソン、ピーター(編)。「世界のトップからの小さな新しいティラノサウルス」。PLoSONE。9(3):e91287。Bibcode:2014PLoSO … 991287F。土井:10.1371 /journal.pone.0091287。PMC 3951350。PMID 24621577。 ^ フィオリロ、アンソニーR。; ガングロフ、ローランドA.(2000)。「アラスカ北部のプリンスクリーク層(白亜紀)からの獣脚類の歯、北極恐竜の古生態学に関する推測」。Journal of VertebratePaleontology。20(4):675 DOI:10.1671 / 0272から4634(2000年)020 2.0.CO; 2。
^ Lü、J .; Yi、L。; ブルサット、SL; ヤン、L。; Chen、L。(2014)「アジアの後期白亜紀の長い鼻のティラノサウルスの新しいクレード」。ネイチャーコミュニケーションズ。5(3788)。Bibcode:2014NatCo … 5.3788L。土井:10.1038 / ncomms4788。PMID 24807588。 ^ ローウェン、マサチューセッツ州; Irmis、RB; Sertich、JJW; カリー、PJ; サンプソン、SD(2013)。エバンス、DC(編)。「暴君恐竜の進化は白亜紀後期の海の上昇と下降を追跡します」。PLoSONE。8(11):e79420。Bibcode:2013PLoSO … 879420L。土井:10.1371 /journal.pone.0079420。PMC 3819173。PMID 24223179。 ^ 徐星、X ; Norell、Mark A。; Kuang Xuewen; 王シャオリン; 趙岐; 嘉成海。(2004)。「中国からの基本的なティラノサウルスとティラノサウルスの原羽の証拠」。ネイチャー。431(7009):680–684。Bibcode:2004Natur.431..680X。土井:10.1038 / nature02855。PMID 15470426。S2CID 4381777。 ^ Xu、X。; 王、K。; 張、K。; Ma、Q。; Xing、L。; サリバン、C。; 胡、D。; チェン、S。; 王、S。; etal。(2012)。「中国の前期白亜紀からの巨大な羽毛恐竜」(PDF)。ネイチャー。484(7392):92–95。土井:10.1038 / nature10906。PMID 22481363。S2CID 29689629。2012年4月17日にオリジナル(PDF)からアーカイブされました。
^ ベル、PR; カンピオーネ、北東; 人、WS; カリー、PJ; ラーソン、PL; タンケ、DH; バッカー、RT(2017)。「ティラノサウルスの外皮は、巨人症と羽毛の進化の相反するパターンを明らかにします」。生物学の手紙。13(6):20170092. DOI:10.1098 / rsbl.2017.0092。PMC 5493735。PMID 28592520。 ^ カー、トーマスD。; Varricchio、David J。; Sedlmayr、Jayc C。; ロバーツ、エリックM。; ムーア、ジェイソンR.(2017-03-30)。「成長不全とワニのような顔の感覚系の証拠を持つ新しいティラノサウルス」。ScientificReports。7:44942. Bibcode:2017NatSR … 744942C。土井:10.1038 / srep44942。ISSN 2045年から2322年。PMC 5372470。PMID 28358353。 ^ ミリンコビッチ、ミシェル; マヌキアン、リアナ; デブリー、エイドリアン; ディポー、ニコラス; マーティン、サミュエル; シン、ダリジット; ランバート、ドミニク; Zwicker、Matthias(2013年1月4日)。「クロコダイルの頭のうろこは発達単位ではありませんが、物理的なひび割れから発生します」。科学。339(6115):78–81。Bibcode:2013Sci … 339 … 78M。土井:10.1126 /science.1226265。PMID 23196908。S2CID 6859452。2020年11月6日にオリジナルからアーカイブされました。
^ lcourt、R。; グリロ、オン(2018)。「南半球からのティラノサウルス:生物地理学、進化、および分類学への影響」。古地理学、古気候学、古生態学。511:379–387。Bibcode:2018PPP … 511..379D。土井:10.1016 /j.palaeo.2018.09.003。S2CID 133830150。 ^ カリー、フィリップJ。; Hurum、JørnH.; サバス、カロル(2003)。「ティラノサウルスの頭蓋骨の構造と進化」(PDF)。Acta PalaeontologicaPolonica。48(2):227–234。2007年10月25日にオリジナル(PDF)からアーカイブされました。
^ Voris、Jared T。; テリエン、フランソワ; Zelenitzky、Darla K。; ブラウン、カレブM.(2020)。「カナダ、アルバータ州のカンパニアフォアモスト累層からの新しいティラノサウルス(獣脚類:ティラノサウルス科)は、ティラノサウルスの進化と生物地理学への洞察を提供します」。白亜紀の研究。110:104388. DOI:10.1016 / j.cretres.2020.104388。
^ ポール、グレゴリーS.(1988)。世界の略奪的な恐竜。ニューヨーク:サイモン&シュスター。頁。 464pp。ISBN 978-0-671-61946-6。
^ ポール、グレゴリーS.(2016)。恐竜へのプリンストンフィールドガイド。プリンストン:プリンストン大学出版局。NS。360. ISBN 9781400883141。”