Wnt_signaling_pathway
Wntシグナル伝達経路はの基であるシグナル伝達で始まる経路タンパク質の信号を通過介して細胞内に細胞表面受容体。Wntという名前は、WinglessとInt-1という名前から作成されたかばん語です。 Wntシグナル伝達経路は、近くの細胞間コミュニケーション(パラクリン)または同じ細胞コミュニケーション(オートクリン)のいずれかを使用します。それらは動物で高度に進化的に保存されています。つまり、ミバエからヒトまで、動物種間で類似しています。
3つのWntシグナル伝達経路が特徴づけられています:標準的なWnt経路、非標準的な平面細胞極性経路、および非標準的なWnt /カルシウム経路。3つの経路はすべて、Wntタンパク質リガンドがFrizzledファミリー受容体に結合することによって活性化されます。Frizzledファミリー受容体は、細胞内のDisheveledタンパク質に生物学的シグナルを渡します。標準的なWnt経路は遺伝子 転写の調節につながり、SPATS1遺伝子によって部分的に負に調節されていると考えられています。非標準的な平面細胞極性経路は、細胞の形状に関与する細胞骨格を調節します。非標準的なWnt /カルシウム経路は細胞内のカルシウムを調節します。
Wntシグナル伝達は、発がんにおけるその役割について最初に特定され、次に胚発生におけるその機能について特定されました。それが制御する胚のプロセスには、体軸のパターン形成、細胞の運命の特定、細胞増殖および細胞移動が含まれます。これらのプロセスは、骨、心臓、筋肉などの重要な組織を適切に形成するために必要です。胚発生におけるその役割は、Wnt経路タンパク質の遺伝子変異が異常なミバエ 胚を生成したときに発見されました。その後の研究では、これらの異常の原因となる遺伝子がマウスの乳がんの発生にも影響を与えることがわかりました。Wntシグナル伝達は、成人の骨髄、皮膚、腸の組織再生も制御します。
この経路の臨床的重要性は、乳がんや前立腺がん、膠芽腫、2型糖尿病などのさまざまな疾患につながる変異によって実証されました。 近年、研究者らは、疾患のマウスモデルにおけるWnt経路阻害剤の最初の使用の成功を報告しました。
コンテンツ
1 歴史と語源
2 タンパク質
3 機構
3.1 財団 3.2 カノニカルおよび非カノニカル経路
3.2.1 カノニカルパスウェイ
3.2.2 非正規経路
3.3 統合されたWnt経路 3.43.4 その他の経路 3.5 規制
4 誘導された細胞応答
4.1 胚発生
4.1.1 軸のパターン化
4.1.2 拘束拘束仕様
4.1.3 細胞増殖
4.1.4 細胞移動
4.2 インスリン感受性
5 臨床的意義
5.1 癌 5.2 II型糖尿病
6 も参照してください
7 参考文献
8 参考文献
9 外部リンク
歴史と語源
Wntシグナル伝達の発見は、発癌性(癌を引き起こす)レトロウイルスの研究によって影響を受けました。1982年に、ロエルNusseとハロルドVarmusはをマウスに感染したマウス乳腺腫瘍ウイルス乳房腫瘍を引き起こす可能性が変異した遺伝子を見るためのmutateマウス遺伝子に順に。彼らは、int1(統合1)と名付けた新しいマウス癌原遺伝子を特定しました。
Int1は、ヒトやショウジョウバエを含む複数の種で高度に保存されています。プレゼンスキイロショウジョウバエにおけるINT1遺伝子ことを導いた研究者が1987年に発見するために、ショウジョウバエは、実際にすでに知られていたと特徴付けショウジョウバエウィングレス(WG)として知られている遺伝子を。 ChristianeNüsslein-VolhardとEricWieschaus(1995年にノーベル生理学・医学賞を受賞)による以前の研究以来、胚発生時の体軸の形成に関与するセグメント極性遺伝子としてのWgの機能はすでに確立されていました。発生、研究者は、マウスで発見された哺乳類のint1が胚発生にも関与していることを確認しました。
継続的な研究により、さらにint1関連遺伝子が発見されました。しかし、これらの遺伝子はint1と同じ方法で同定されなかったため、int遺伝子の命名法は不十分でした。したがって、int / WinglessファミリーはWntファミリーになり、int1はWnt1になりました。Wntという名前はintとWgのかばん語で、「Wingless関連の統合サイト」の略です。
タンパク質

Wnt8の結晶タンパク質構造とFrizzled8のシステインリッチドメイン
Wntは、350〜400アミノ酸の長さの分泌脂質修飾シグナル伝達糖タンパク質の多様なファミリーで構成されています。すべてのWntの脂質修飾はpalmitoleoylation単一の完全に保存されたセリン残基の。パルミトレオイル化が必要なのは、Wntがそのキャリアタンパク質Wntless(WLS)に結合して分泌のために原形質膜に輸送され、Wntタンパク質がその受容体Frizzledに結合できるようにするためです。 Wntタンパク質はまた、適切な分泌を確実にするために炭水化物を結合するグリコシル化を受けます。 Wntシグナル伝達では、これらのタンパク質は、パラクリンおよびオートクリン経路を介して異なるWnt経路を活性化するリガンドとして機能します。
これらのタンパク質は、種間で高度に保存されています。それらは、マウス、ヒト、アフリカツメガエル、ゼブラフィッシュ、ショウジョウバエなどに見られます。
種族
Wntタンパク質
ホモサピエンス
WNT1、WNT2、WNT2B、WNT3、WNT3A、WNT4、WNT5A、WNT5B、WNT6、WNT7A、WNT7B、WNT8A、WNT8B、WNT9A、WNT9B、WNT10A、Wnt10bで、WNT11、WNT16
ハツカネズミ(H. sapiensと同じタンパク質)
Wnt1、Wnt2、Wnt2B、Wnt3、Wnt3A、Wnt4、Wnt5A、Wnt5B、Wnt6、Wnt7A、Wnt7B、Wnt8A、Wnt8B、Wnt9A、Wnt9B、Wnt10A、Wnt10B、Wnt11、Wnt16 Xenopus Wnt1、Wnt2、Wnt2B、Wnt3、Wnt3A、Wnt4、Wnt5A、Wnt5B、Wnt7A、Wnt7B、Wnt8A、Wnt8B、Wnt10A、Wnt10B、Wnt11、Wnt11R
ダニオ・レリオ
Wnt1、Wnt2、Wnt2B、Wnt3、Wnt3A、Wnt4、Wnt5A、Wnt5B、Wnt6、Wnt7A、Wnt7B、Wnt8A、Wnt8B、Wnt10A、Wnt10B、Wnt11、Wnt16
ショウジョウバエ
Wg、DWnt2、DWnt3 / 5、DWnt 4、DWnt6、WntD / DWnt8、DWnt10
ヒドラ
hywnt1、hywnt5a、hywnt8、hywnt7、hywnt9 / 10a、hywnt9 / 10b、hywnt9 / 10c、hywnt11、hywnt16
C.エレガンス
mom-2、lin-44、egl-20、cwn-1、cwn-2
機構

図2.Wntは受容体に結合(活性化)します。アキシンは「破壊複合体」から削除されます。β-Catは核内に移動し、DNAの転写因子に結合し、タンパク質の転写を活性化します。「P」はリン酸塩を表し 
図1.Wntは受容体に結合しません。Axin、GSK、APCは「破壊複合体」を形成し、β-Catは破壊されます。
財団
Wntシグナル伝達は、Wntタンパク質がFrizzled(Fz)ファミリー受容体のN末端細胞外システインリッチドメインに結合したときに始まります。これらの受容体は原形質膜に7回広がり、Gタンパク質共役型受容体(GPCR)の異なるファミリーを構成します。しかしながら、Wntシグナル伝達を促進するために、Wntタンパク質とFz受容体の間の相互作用とともに補助受容体が必要となる場合が例としては、リポタンパク質受容体関連タンパク質(LRP)-5/6、受容体型チロシンキナーゼ(RTK)、ROR2などが受容体が活性化されると、細胞質にあるリンタンパク質 Disheveled(Dsh)に信号が送られます。この信号は、FzとDshの間の直接的な相互作用を介して送信されます。Dshタンパク質はすべての生物に存在し、それらはすべて次の高度に保存されたタンパク質ドメインを共有しています:アミノ末端DIXドメイン、中央PDZドメイン、およびカルボキシ末端DEPドメイン。Dshの後、Wntシグナルは複数の経路に分岐し、各経路は3つのドメインの異なる組み合わせと相互作用するため、これらの異なるドメインは重要です。
カノニカルおよび非カノニカル経路
3つの最も特徴的なWntシグナル伝達経路は、標準的なWnt経路、非標準的な平面細胞極性経路、および非標準的なWnt /カルシウム経路です。それらの名前が示すように、これらの経路は、正規または非正規の2つのカテゴリのいずれかに属します。カテゴリ間の違いは、標準的な経路にはタンパク質β-カテニンが含まれるのに対し、非標準的な経路はそれとは独立して機能することです。

標準的なWnt経路
カノニカルパスウェイ
カノニカルWnt経路(またはWnt /βカテニン経路)の蓄積させるWnt経路であるβカテニンに細胞質におけるその最終的な転位を核転写として作用するコアクチベーターの転写因子に属するTCF / LEFファミリー。Wntがないと、破壊複合体が通常細胞質を分解するため、β-カテニンは細胞質に蓄積しません。この破壊複合体には、Axin、大腸腺腫症(APC)、プロテインホスファターゼ2A(PP2A)、グリコーゲンシンターゼキナーゼ3(GSK3)、カゼインキナーゼ1α(CK1α)のタンパク質が含まれています。 それは、ユビキチン化の標的となることによってβ-カテニンを分解し、その後、それをプロテアソームに送って消化します。 しかし、WntがFzとLRP5 / 6に結合するとすぐに、破壊複合体機能が破壊されます。これは、Wntが負のWntレギュレーターであるAxinの移行を引き起こし、破壊複合体が原形質膜に移動するためです。破壊複合体の他のタンパク質によるリン酸化は、その後、AxinをLRP5 / 6の細胞質尾部に結合します。アキシンは脱リン酸化され、その安定性とレベルが低下します。その後、Dshはリン酸化を介して活性化され、そのDIXおよびPDZドメインは破壊複合体のGSK3活性を阻害します。これにより、β-カテニンが蓄積して核に局在し、その後、TCF / LEF(T細胞因子/リンパ球増強因子)転写因子とともに遺伝子導入を介して細胞応答を誘導することができます。 β-カテニンは、BCL9、Pygopus 、パラフィブロミン/イワダヌキなどの他の転写コアクチベーターを動員します。新しいハイスループットプロテオミクス研究のおかげで、β-カテニンによって組み立てられた転写複合体の複雑さが明らかになり始めています。しかしながら、β-カテニンがどのように標的遺伝子の発現を促進するかについての統一理論はまだ見当たらず、組織特異的なプレーヤーはβ-カテニンがその標的遺伝子を定義するのを助けるかもしれません。 β-カテニン相互作用タンパク質の拡張性は私たちの理解を複雑にします:β-カテニンはAktによってSer552で直接リン酸化される可能性があり、それが細胞間接触からの分離と細胞質ゾルへの蓄積を引き起こし、その後14-3-3ζが相互作用しますβ-カテニン(pSer552)とその核転座を強化します。 BCL9とPygopusは、実際、いくつかのβ-カテニン非依存性機能を持っていると報告されています(したがって、おそらくWntシグナル伝達非依存性)。 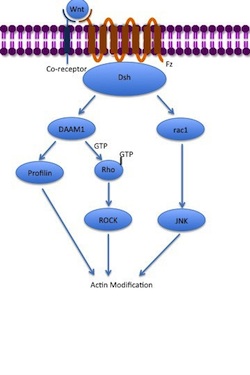
非正規PCP経路
非正規経路
非標準的な平面細胞極性(PCP)経路には、β-カテニンは含まれません。コレセプターとしてLRP-5 / 6を使用せず、NRH1、Ryk、PTK7、またはROR2を使用すると考えられています。PCP経路は、WntがFzとその補助受容体に結合することで活性化されます。次に、受容体はDshを動員し、DshはそのPDZおよびDIXドメインを使用して、形態形成のDishevelled関連活性化因子1(DAAM1)と複合体を形成します。Daam1は、グアニン交換因子を介して小さなGタンパク質 Rhoを活性化します。Rhoは、細胞骨格の主要な調節因子の1つであるRho関連キナーゼ(ROCK)を活性化します。Dshはまた、rac1と複合体を形成し、アクチンへのプロフィリンの結合を仲介します。Rac1はJNKを活性化し、アクチン重合を引き起こす可能性もプロフィは、アクチンに結合して細胞骨格の再編につながることができ、原腸陥入。 
非標準的なWnt /カルシウム経路
非標準的なWnt /カルシウム経路もβ-カテニンを含みません。その役割は、細胞内カルシウムレベルを制御するために、小胞体(ER)からのカルシウム放出の調節を助けることです。他のWnt経路と同様に、リガンドが結合すると、活性化されたFz受容体はDshと直接相互作用し、特定のDshタンパク質ドメインを活性化します。Wnt /カルシウムシグナル伝達に関与するドメインは、PDZドメインとDEPドメインです。ただし、他のWnt経路とは異なり、Fz受容体は三量体Gタンパク質と直接相互作用します。DshとGタンパク質のこの共刺激は、PLCまたはcGMP固有のPDEのいずれかの活性化につながる可能性がPLCがアクティブになると、原形質膜コンポーネントPIP2がDAGとIP3に切断されます。IP3がER上の受容体に結合すると、カルシウムが放出されます。カルシウムとDAGの濃度が高くなると、PKCを介してCdc42が活性化される可能性がCdc42は、腹側パターン形成の重要な調節因子です。カルシウムの増加は、カルシニューリンとCaMKIIも活性化します。CaMKIIは、細胞接着、遊走、組織分離を調節する転写因子NFATの活性化を誘導します。カルシニューリンは、TAK1およびNLKキナーゼを活性化します。これらは、標準的なWnt経路におけるTCF /β-カテニンシグナル伝達を妨害する可能性がしかしながら、PDEが活性化されると、ERからのカルシウム放出が阻害されます。PDEは、PKGの阻害を介してこれを媒介し、これは、その後、カルシウム放出の阻害を引き起こす。
統合されたWnt経路
カノニカルおよび非カノニカルWntシグナル伝達経路の二元的な区別が精査されており、統合された収束Wnt経路が提案されています。これに関するいくつかの証拠が1つのWntリガンド(Wnt5A)で発見されました。複数のWntリガンドについて、Wnt / Ca2 +およびWnt /β-カテニンシグナル伝達の統合された活性化を示す収束Wntシグナル伝達経路の証拠が哺乳類細胞株で報告された。
その他の経路
Wntシグナル伝達は、それほど広く解明されていない他の多くのシグナル伝達経路も調節します。そのような経路の1つには、WntとGSK3の間の相互作用が含まれます。細胞増殖中、Wntはβ-カテニンの非存在下でmTORを活性化するためにGSK3を阻害することができます。ただし、Wntは、DshとGSK3の相互作用を介してアップレギュレートされる腫瘍抑制因子 TSC2の活性化を介して、mTORの負のレギュレーターとしても機能します。筋形成の間、WntはPAおよびCREBを使用してMyoDおよびMyf5遺伝子を活性化します。 Wntはまた、RykおよびSrcと連携して作用し、軸索ガイダンス中のニューロンの反発の調節を可能にします。CK1が原腸陥入時に細胞骨格を調節するためにRap1-ATPaseの阻害剤として機能する場合、Wntは原腸陥入を調節します。Wntが一緒にROR2を使用する場合、原腸形成のさらなる調節が達成され、CDC42及びJNKの発現を調節する経路PAPCを。Dshは、細胞極性と微小管細胞骨格の発達を制御するために、aPKC、Pa3、Par6、およびLG1と相互作用することもできます。これらの経路は、PCPおよびWnt /カルシウムシグナル伝達に関連する成分と重複していますが、異なる応答を生成するため、別個の経路と見なされます。
規制
適切な機能を確保するために、Wntシグナル伝達はそのシグナル伝達経路に沿ったいくつかのポイントで絶えず調節されています。たとえば、Wntタンパク質はパルミトイル化されています。タンパク質ヤマアラシはこのプロセスを仲介します。つまり、Wntリガンドが完全に形成される時期を決定することにより、いつ分泌されるかを調節するのに役立ちます。分泌は、GPR177(wntless)などのタンパク質、中断された均一性、レトロマー複合体などの複合体によってさらに制御されます。
時分泌、リガンドは、安定剤としてのタンパク質の結合を介してその受容体に到達することを防止することができるダリーとグリピカン3(GPC3)、禁止拡散。癌細胞では、GPC3のヘパラン硫酸鎖 とコアタンパク質 の両方が、細胞増殖のためのWnt結合と活性化の調節に関与しています。 Wntは、IdoA2SとGlcNS6Sを含むGPC3のヘパラン硫酸構造を認識し、GlcNS6S3Sの3-O-硫酸化は、ヘパラン硫酸グリピカンへのWntの結合を強化します。 GPC3のNローブにあるシステインに富むドメインは、Wntと相互作用するフェニルアラニン-41を含むWnt結合疎水性溝を形成することが確認されています。 HN3と呼ばれるナノボディを使用してWnt結合ドメインをブロックすると、Wntの活性化を阻害する可能性が
Fz受容体では、Wnt以外のタンパク質の結合がシグナル伝達に拮抗する可能性が具体的なアンタゴニストには含まDickkopf(のDkk)、wnt阻害因子1(WIF-1)、 、分泌frizzled関連タンパク質(SFRP)、ケルベロス、FRZB、ワイズ、SOST、及びネイキッドキューティクル。これらはWntシグナル伝達の阻害剤を構成します。ただし、他の分子も活性化因子として機能します。NorrinとR-Spondin2は、Wntリガンドの非存在下でWntシグナル伝達を活性化します。
Wntシグナル伝達経路間の相互作用もWntシグナル伝達を調節します。前述のように、Wnt /カルシウム経路はTCF /β-カテニンを阻害し、標準的なWnt経路のシグナル伝達を妨げる可能性が プロスタグランジンE2は、標準的なWntシグナル伝達経路の必須の活性化因子です。PGE2とその受容体E2 / E4との相互作用は、cAMP / PKAを介したリン酸化を介してβ-カテニンを安定化します。PGE2の合成は、ゼブラフィッシュやマウスの組織再生や幹細胞集団の制御など、Wntシグナル伝達を介したプロセスに必要です。興味深いことに、いくつかの特大の天然変性タンパク質の非構造化領域は、Wntシグナル伝達の調節に重要な役割を果たしています。
誘導された細胞応答編集
胚発生
Wntシグナル伝達は、胚発生において重要な役割を果たします。脊椎動物と無脊椎動物の両方で機能します。これには、人間、カエル、ゼブラフィッシュ、線虫、ショウジョウバエなどが含まれます。それはショウジョウバエのセグメント極性で最初に発見されました、そこでそれは前部と後部の極性を確立するのを助けます。それは他の発達過程に関係しています。ショウジョウバエでのその機能が示唆するように、それは体軸形成、特に前後軸と背腹軸の形成において重要な役割を果たします。細胞分化の誘導に関与し、肺や卵巣などの重要な臓器の形成を促します。Wntはさらに、細胞の増殖と移動を適切に制御することにより、これらの組織の発達を確実にします。Wntシグナル伝達機能は、軸のパターン形成、細胞の運命の特定、細胞増殖、細胞移動に分けることができます。
軸のパターン化
初期の胚の発達において、一次体軸の形成は、生物の全体的な体の計画を確立する上で重要なステップです。軸には、前後軸、背腹軸、および左右軸が含まれます。Wntシグナル伝達は、前後軸と背腹(DV)軸の形成に関係しています。前後の発達におけるWntシグナル伝達活性は、哺乳類、魚、カエルに見られます。哺乳類では、原始線条および他の周囲の組織が形態形成化合物Wnt、BMP、FGF、ノーダルおよびレチノイン酸を産生して、原腸陥入後期に後部領域を確立します。これらのタンパク質は濃度勾配を形成します。最高濃度の領域は後部領域を確立し、最低濃度の領域は前部領域を示します。魚やカエルでは、標準的なWntシグナル伝達によって生成されたβ-カテニンが組織化センターの形成を引き起こし、BMPと並んで後部形成を誘発します。DV軸形成へのWntの関与は、背側領域を確立するシュペーマンオーガナイザーの形成の活動に見ることができます。標準的なWntシグナル伝達β-カテニン産生は、双子とシャムの遺伝子の活性化を介してこのオーガナイザーの形成を誘導します。 同様に、鳥類の原腸陥入では、コラーの鎌の細胞は、原始線条の形成中に細胞の異なる動きを可能にする異なる中胚葉マーカー遺伝子を発現します。FGFによって活性化されるWntシグナル伝達がこの動きの原因です。
Wntシグナル伝達は、開発の後半にある特定の体の部分や臓器系の軸形成にも関与しています。脊椎動物では、ソニックヘッジホッグ(Shh)とWnt形態形成シグナル伝達勾配が、神経管の軸方向のパターン形成中に中枢神経系の背腹軸を確立します。高Wntシグナル伝達は背側領域を確立し、高Shhシグナル伝達は腹側領域を示します。 Wntは、軸索ガイダンスへの関与を通じて中枢神経系のDV形成に関与しています。Wntタンパク質は、脊髄の軸索を前後方向に誘導します。 Wntは四肢のDV軸の形成にも関与しています。具体的には、Wnt7aは、発達中の四肢の背側のパターンを生成するのに役立ちます。
では胚分化波の開発のWntのモデルを区別するために準備ができたコンピテント細胞にシグナル伝達複合体の一部として重要な役割を果たしています。Wntは細胞骨格の活動に反応し、収縮または拡大の通過波によって作成された初期変化を安定化すると同時に、個々の細胞がどの波に参加したかに関する異なるシグナル伝達経路を使用して核にシグナルを送ります。開発中に発生する機械的シグナル伝達。
拘束拘束仕様
細胞運命の特定または細胞分化は、未分化細胞がより特殊な細胞型になる可能性があるプロセスです。Wntシグナル伝達は、多能性幹細胞の中胚葉および内胚葉 前駆細胞への分化を誘導します。これらの前駆細胞はさらに、内皮、心臓、血管の平滑筋系統などの細胞型に分化します。 Wntシグナル伝達は、幹細胞からの血液形成を誘導します。具体的には、Wnt3は造血能を持つ中胚葉コミット細胞につながります。 Wnt1は神経分化に拮抗し、神経幹細胞の自己複製の主要な要因です。これにより、神経系細胞の再生が可能になります。これは、神経幹細胞の増殖を促進する役割のさらなる証拠です。 Wntシグナル伝達は、生殖細胞の決定、腸組織の特定、毛包の発達、肺組織の発達、体幹神経堤細胞の分化、腎の発達、卵巣の発達、および性別の決定に関与しています。 Wntシグナル伝達も心臓形成に拮抗し、Wnt阻害は発達中の心臓組織の重要な誘導因子であることが示され 、小分子Wnt阻害剤は多能性幹細胞から心筋細胞を産生するために日常的に使用されています。
細胞増殖
異なる生物の特定の細胞組織を形成するために必要な細胞の大量分化を行うために、胚性幹細胞の増殖と成長が起こらなければなりません。このプロセスは、核および細胞質のβ-カテニンを増加させる標準的なWntシグナル伝達によって媒介されます。β-カテニンの増加は、細胞周期におけるG1からSへの相転移を制御するサイクリンD1やc-mycなどのタンパク質の転写活性化を開始する可能性がS期に入ると、DNA複製が起こり、最終的には有糸分裂が起こり、細胞増殖の原因となります。幹細胞が増殖すると、幹細胞も分化するため、この増殖の増加は細胞分化と直接対になっています。これにより、胚発生中の特定の組織システムの全体的な成長と発達が可能になります。これは、Wnt3aが赤血球形成に必要な造血幹細胞の増殖と拡大をもたらす循環器系などのシステムで明らかです。
がん幹細胞の生化学は、他の腫瘍細胞の生化学とは微妙に異なります。これらのいわゆるWnt依存細胞はハイジャックし、Wnt経路の絶え間ない刺激に依存して、制御されていない成長、生存、および移動を促進します。癌、Wntシグナル伝達は、正常な受容体が信号を受信していないにもかかわらず、恒久的に活性化になる下流癌遺伝子および腫瘍抑制遺伝子における突然変異を介して、定期的な刺激とは無関係になることができます。β-カテニンはタンパク質TCF4などの転写因子に結合し、組み合わせて分子が必要な遺伝子を活性化します。LF3は、in vitro、細胞株でこの結合を強力に阻害し、マウスモデルで腫瘍増殖を抑制します。それは複製を防ぎ、移動する能力を低下させましたが、すべて健康な細胞に影響を与えることはありませんでした。治療後、がん幹細胞は残っていませんでした。この発見は、AlphaScreensとELISAテクノロジーを含む「合理的なドラッグデザイン」の成果でした。
細胞移動

上皮間葉転換を示す図
胚発生中の細胞移動は、体軸の確立、組織形成、四肢の誘導、および他のいくつかのプロセスを可能にします。Wntシグナル伝達は、特に収束伸長中に、このプロセスを仲介するのに役立ちます。原腸陥入時の適切な収束伸長には、WntPCP経路と標準的なWnt経路の両方からのシグナル伝達が必要です。収束伸長は、活性化されると収束伸長をブロックするWnt /カルシウム経路によってさらに調節されます。Wntシグナル伝達はまた、神経芽細胞、神経堤細胞、筋細胞、および気管細胞の遊走挙動の制御を通じて、発生の後期段階で細胞遊走を誘導します。
Wntシグナル伝達は、上皮間葉転換(EMT)として知られる別の重要な移行プロセスに関与しています。このプロセスにより、上皮細胞が間葉系細胞に変化し、ラミニンで所定の位置に保持されなくなります。これには、細胞がラミニンから分離して移動できるように、カドヘリンのダウンレギュレーションが含まれます。Wntシグナル伝達は、特に乳腺の発達において、EMTの誘導因子です。
インスリン感受性

Wntシグナル伝達経路とインスリンシグナル伝達経路の間の相互作用を示す図
インスリンは、特定の生物の血糖恒常性に関与するペプチドホルモンです。具体的には、血流からのグルコース取り込みを増加させるために、細胞膜のグルコーストランスポーターのアップレギュレーションにつながります。このプロセスは、Wnt /β-カテニンシグナル伝達の活性化によって部分的に媒介され、細胞のインスリン感受性を高める可能性が特に、Wnt10bは骨格筋細胞のこの感受性を高めるWntタンパク質です。
臨床的意義編集
癌
その最初の発見以来、Wntシグナル伝達は癌と関連がありました。Wnt1が発見されたとき、それは最初のプロトとして同定された癌遺伝子でマウスモデル乳がんのため。Wnt1がWgのホモログであるという事実は、それが胚発生に関与していることを示しており、これはしばしば迅速な細胞分裂と移動を必要とします。これらのプロセスの誤調節は、過剰な細胞増殖を介して腫瘍の発生につながる可能性が
標準的なWnt経路の活動は、良性および悪性の乳房腫瘍の発生に関与しています。腫瘍の化学療法抵抗性におけるWnt経路の役割、および癌開始細胞の明確な亜集団の維持におけるその役割も十分に文書化されています。その存在は、核および/または細胞質におけるβ-カテニンのレベルの上昇によって明らかになり、免疫組織化学的染色およびウエスタンブロッティングで検出することができます。β-カテニン発現の増加は、乳がん患者の予後不良と相関しています。この蓄積は、β-カテニンの変異、β-カテニン破壊複合体の欠損、最も頻繁にはAPCの構造的に無秩序な領域の変異、Wntリガンドの過剰発現、阻害剤の喪失、および/または調節活性の低下などの要因による可能性があります経路(Wnt /カルシウム経路など)。 乳房腫瘍は、EMTへのWntの関与により転移する可能性がある。基底細胞様乳がんの肺への転移を調べた研究では、Wnt /β-カテニンシグナル伝達の抑制により、転移を阻害する可能性のあるEMTを防ぐことができることが示されました。
Wntシグナル伝達は、他の癌の発症に関係しています。変更CTNNB1の遺伝子エンコードβカテニンつまり式は、胸に測定することができ、大腸、黒色腫、前立腺癌、肺癌、および他の癌。Wnt1、Wnt2、Wnt7AなどのWntリガンドタンパク質の発現増加は、それぞれ膠芽腫、食道癌、卵巣癌の発症で観察されました。適切に機能しない場合に複数の種類の癌を引き起こす他のタンパク質には、ROR1、ROR2、SFRP4、Wnt5A、WIF1、およびTCF / LEFファミリーのタンパク質が含まれます。
PGE2とWntの関連性は、PGE2の慢性的な炎症関連の増加が、さまざまな組織でWnt経路の活性化を引き起こし、発がんを引き起こす可能性があることを示唆しています。
II型糖尿病
2型糖尿病は、末梢でのインスリン分泌の低下とインスリン抵抗性の増加を引き起こす一般的な疾患です。その結果、血糖値が上昇したり、高血糖が発生したりします。これは、治療しないと致命的となる可能性がWntシグナル伝達はインスリン感受性に関与しているため、その経路の機能不全が関与している可能性がたとえば、Wnt5bの過剰発現は、肥満と2型糖尿病の併存疾患が高いため、脂肪生成におけるその役割のために感受性を高める可能性が Wntシグナル伝達は、ミトコンドリア生合成の強力な活性化因子です。これにより、DNAや細胞の損傷を引き起こすことが知られている活性酸素種(ROS)の生成が増加します。このROS誘発性損傷は、急性肝インスリン抵抗性または傷害誘発性インスリン抵抗性を引き起こす可能性があるため、重大です。 TCF7L2などのWntシグナル伝達関連転写因子の変異は、感受性の増加に関連しています。
も参照してくださいAXIN1 GSK-3
脱毛の管理
ウィングレスローカリゼーションエレメント3(WLE3)
WNT1誘導性シグナル伝達経路タンパク質1(WISP1)
WNT1誘導性シグナル伝達経路タンパク質2(WISP2)
WNT1誘導性シグナル伝達経路タンパク質3(WISP3)
参考文献
^ Nusse R、Brown A、Papkoff J、Scambler P、Shackleford G、McMahon A、他 (1991年1月)。「int-1および関連遺伝子の新しい命名法:Wnt遺伝子ファミリー」。セル。64(2):231 DOI:10.1016 / 0092から8674(91)、90633。PMID 1846319。S2CID 3189574。
^ Nusse R、Varmus HE(1992年6月)。「Wnt遺伝子」。セル。69(7):1073–87。土井:10.1016 / 0092-8674(92)90630-U。PMID 1617723。
^ Nusse R。「疾患および発生中のWntシグナル伝達」。セルリサーチ。15(1):28–32。土井:10.1038 /sj.cr.7290260。PMID 15686623。
^ Zhang H、Zhang H、Zhang Y、Ng SS、Ren F、Wang Y、Duan Y、Chen L、Zhai Y、Guo Q、Chang Z。「Dishevelled-DEPドメイン相互作用タンパク質(DDIP)は、TCF4分解を促進し、TCF4 /β-カテニン複合体を破壊することにより、Wntシグナル伝達を阻害します」。細胞シグナル伝達。22(11):1753–60。土井:10.1016 /j.cellsig.2010.06.016。PMID 20603214。
^ Goessling W、North TE、Loewer S、Lord AM、Lee S、Stoick-Cooper CL、Weidinger G、Puder M、Daley GQ、Moon RT、Zon LI。「PGE2とWntシグナル伝達の遺伝的相互作用は、幹細胞の発生仕様と再生を調節します」。セル。136(6):1136–47。土井:10.1016 /j.cell.2009.01.015。PMC 2692708。PMID 19303855。
^ Logan CY、Nusse R(2004)。「発生と疾患におけるWntシグナル伝達経路」。細胞および発生生物学の年次レビュー。20:781〜810。CiteSeerX 10.1.1.322.311。土井:10.1146 /annurev.cellbio.20.010403.113126。PMID 15473860。
^ Komiya Y、Habas R。「Wntシグナル伝達経路」。器官形成。4(2):68–75。土井:10.4161 /org.4.2.5851。PMC 2634250。PMID 19279717。
^ Zimmerli D、Hausmann G、CantùC、Basler K。「Wnt経路への薬理学的介入:核内因子のタンパク質間相互作用の破壊に対するWnt分泌の阻害」。ブリティッシュジャーナルオブファーマコロジー。174(24):4600–4610。土井:10.1111 /bph.13864。PMC 5727313。PMID 28521071。
^ Nusse R、van Ooyen A、Cox D、Fung YK、Varmus H(1984)。「マウス15番染色体上の推定乳腺癌遺伝子(int-1)のプロウイルス活性化のモード」。ネイチャー。307(5947):131–6。Bibcode:1984Natur.307..131N。土井:10.1038 / 307131a0。PMID 6318122。S2CID 4261052。
^ Klaus A、Birchmeier W。「Wntシグナル伝達とその発生および癌への影響」。ネイチャーレビュー。がん。8(5):387–98。土井:10.1038 / nrc2389。PMID 18432252。S2CID 31382024。
^ Cadigan KM、Nusse R(1997年12月)。「Wntシグナル伝達:動物の発達における共通のテーマ」。遺伝子と開発。11(24):3286–305。土井:10.1101 /gad.11.24.3286。PMID 9407023。
^ Hannoush RN。「合成タンパク質脂質化」。化学生物学における現在の意見。28:39–46。土井:10.1016 /j.cbpa.2015.05.025。PMID 26080277。
^ Yu J、Chia J、Canning CA、Jones CM、Bard FA、Virshup DM。「Wnt分泌中の小胞体へのWLS逆行性輸送」。発生細胞。29(3):277–91。土井:10.1016 /j.devcel.2014.03.016。PMID 24768165。
^ Janda CY、Waghray D、Levin AM、Thomas C、Garcia KC。「FrizzledによるWnt認識の構造的基礎」。科学。337(6090):59–64。Bibcode:2012Sci … 337 … 59J。土井:10.1126 /science.1222879。PMC 3577348。PMID 22653731。
^ Hosseini V、Dani C、Geranmayeh MH、Mohammadzadeh F、Nazari Soltan Ahmad S、Darabi M。「Wnt脂質化:人身売買、調節、および機能における役割」。Journal of CellularPhysiology。234(6):8040–8054。土井:10.1002 /jcp.27570。PMID 30341908。S2CID 53009014。
^ 倉吉M、山本H、泉S、菊池A。「Wnt-5aの翻訳後パルミトイル化とグリコシル化は、そのシグナル伝達に必要です」。生化学ジャーナル。402(3):515–23。土井:10.1042 / BJ20061476。PMC 1863570。PMID 17117926。
^ ヌッセ、ロエル。「Wntホームページ」。取得した15年4月2013。
^ Sawa H、Korswagen HC。「C.エレガンスにおけるWNTシグナル伝達」。WormBook:1〜30。土井:10.1895 /wormbook.1.7.2。PMC 5402212。PMID 25263666。
^ Rao TP、KühlM。「Wntシグナル伝達経路に関する最新の概要:詳細の前置き」。循環研究。106(12):1798–806。土井:10.1161 /CIRCRESAHA.110.219840。PMID 20576942。
^ Schulte G、Bryja V。「型破りなGタンパク質共役型受容体のFrizzledファミリー」。薬理学の動向。28(10):518–25。土井:10.1016 /j.tips.2007.09.001。PMID 17884187。
^ Habas R、Dawid IB。「乱れたWntシグナル伝達:核は最後のフロンティアですか?」。Journal ofBiology。4(1):2。DOI:10.1186 / jbiol22。PMC 551522。PMID 15720723。
^ Minde DP、Anvarian Z、RüdigerSG、Maurice MM。「混乱の障害:腫瘍抑制タンパク質APCのミスセンス変異はどのようにして癌につながるのか?」。分子がん。10:101 DOI:10.1186 / 1476-4598-10-101。PMC 3170638。PMID 21859464。
^ Minde DP、Radli M、Forneris F、Maurice MM、RüdigerSG(2013)。バックルAM(編)。「腺腫性ポリポーシスコリの大規模な障害は、点突然変異からWntシグナル伝達を保護する戦略を提供します」。PLOSONE。8(10):e77257。Bibcode:2013PLoSO … 877257M。土井:10.1371 /journal.pone.0077257。PMC 3793970。PMID 24130866。
^ MacDonald BT、Tamai K、He X。「Wnt /ベータカテニンシグナル伝達:成分、メカニズム、および疾患」。発生細胞。17(1):9–26。土井:10.1016 /j.devcel.2009.06.016。PMC 2861485。PMID 19619488。
^ Staal FJ、Clevers H。「T細胞発達中のTcf / Lef転写因子:ユニークで重複する機能」。血液学ジャーナル。1(1):3–6。土井:10.1038 /sj.thj.6200001。PMID 11920163。
^ Kramps T、Peter O、Brunner E、Nellen D、Froesch B、Chatterjee S、Murone M、ZülligS、Basler K。「Wnt /ウィングレスシグナル伝達には、BCL9 /レッグレスを介した核ベータ-カテニン-TCF複合体へのpygopusの動員が必要です」(PDF)。セル。109(1):47–60。土井:10.1016 / s0092-8674(02)00679-7。PMID 11955446。S2CID 16720801。
^ Mosimann C、Hausmann G、Basler K。「パラフィブロミン/イワダヌキは、ベータカテニン/アルマジロと直接結合することにより、Wnt / Wg標的遺伝子の転写を活性化します」。セル。125(2):327–41。土井:10.1016 /j.cell.2006.01.053。PMID 16630820。
^ van Tienen LM、Mieszczanek J、Fiedler M、Rutherford TJ、Bienz M。「Legless / BCL9による複数のWntエンハンセオソーム成分の構成的足場」。eLife。6:e20882。土井:10.7554 /elife.20882。PMC 5352222。PMID 28296634。
^ Söderholm、Simon; カントゥ、クラウディオ(2020年10月21日)。「WNT /β-カテニン依存性転写:組織特異的ビジネス」。ワイヤーシステム生物学と医学。13(3):e1511。土井:10.1002 /wsbm.1511。PMID 33085215。
^ Fang D、Hawke D、Zheng Y、Xia Y、Meisenhelder J、Nika H、Mills GB、Kobayashi R、Hunter T、Lu Z。「AKTによるベータカテニンのリン酸化は、ベータカテニンの転写活性を促進します」。Journal of BiologicalChemistry。282(15):11221–9。土井:10.1074 /jbc.M611871200。PMC 1850976。PMID 17287208。
^ CantùC、Valenta T、Hausmann G、Vilain N、Aguet M、Basler K。「Pygo2-H3K4me2 / 3の相互作用は、マウスの発達とWntシグナル伝達に依存する転写に不可欠です」。開発。140(11):2377–86。土井:10.1242 /dev.093591。PMID 23637336。
^ CantùC、Zimmerli D、Hausmann G、Valenta T、Moor A、Aguet M、Basler K。「マウス水晶体発達におけるBcl9タンパク質のPax6依存性であるがβ-カテニン非依存性の機能」。遺伝子と開発。28(17):1879–84。土井:10.1101 /gad.246140.114。PMC 4197948。PMID 25184676。
^ CantùC、Pagella P、Shajiei TD、Zimmerli D、Valenta T、Hausmann G、Basler K、Mitsiadis TA。「歯のエナメル質形成におけるWnt /β-カテニン転写補因子Bcl9、Bcl9l、およびPygopusの細胞質の役割」。科学シグナリング。10(465):eaah4598。土井:10.1126 /scisignal.aah4598。PMID 28174279。S2CID 6845295。
^ Gordon MD、Nusse R。「Wntシグナル伝達:複数の経路、複数の受容体、および複数の転写因子」。Journal of BiologicalChemistry。281(32):22429–33。土井:10.1074 /jbc.R600015200。PMID 16793760。
^ 杉村R、李L。「脊椎動物の発達、幹細胞、および疾患における非標準的なWntシグナル伝達」。先天性欠損症の研究。パートC、今日の胚。90(4):243–56。土井:10.1002 /bdrc.20195。PMID 21181886。
^ van Amerongen R、Nusse R。「開発中のWntシグナル伝達の統合ビューに向けて」。開発。136(19):3205–14。土井:10.1242 /dev.033910。PMID 19736321。
^ van Amerongen R、Fuerer C、Mizutani M、Nusse R。「Wnt5aは、マウス胚発生時にWnt /β-カテニンシグナル伝達を活性化および抑制することができます」。発生生物学。369(1):101–14。土井:10.1016 /j.ydbio.2012.06.020。PMC 3435145。PMID 22771246。
^ Thrasivoulou C、Millar M、Ahmed A。「複数のWntリガンドによる細胞内カルシウムの活性化とβ-カテニンの核への移行:Wnt / Ca2 +およびWnt /β-カテニン経路の収束モデル」。Journal of BiologicalChemistry。288(50):35651–9。土井:10.1074 /jbc.M112.437913。PMC 3861617。PMID 24158438。
^ Inoki K、Ouyang H、Zhu T、Lindvall C、Wang Y、Zhang X、Yang Q、Bennett C、Harada Y、Stankunas K、Wang CY、He X、MacDougald OA、You M、Williams BO、Guan KL(9月2006)。「TSC2は、AMPKとGSK3による協調的なリン酸化を介してWntシグナルとエネルギーシグナルを統合し、細胞増殖を調節します」。セル。126(5):955–68。土井:10.1016 /j.cell.2006.06.055。PMID 16959574。S2CID 16047397。
^ Kuroda K、Kuang S、Taketo MM、Rudnicki MA。「標準的なWntシグナル伝達は、BMP-4が胎児の筋芽細胞の遅い筋形成を特定するように誘導します」。骨格筋。3(1):5。DOI:10.1186 / 2044-5040-3-5。PMC 3602004。PMID 23497616。
^ Malinauskas T、Jones EY。「Wntシグナル伝達の細胞外モジュレーター」。構造生物学における現在の意見。29:77–84。土井:10.1016 /j.sbi.2014.10.003。PMID 25460271。
^ Gao W、Kim H、Feng M、Phung Y、Xavier CP、Rubin JS、Ho M。「肝臓癌治療のためのグリピカン-3のヘパラン硫酸鎖を認識するヒト抗体によるWntシグナル伝達の不活性化」。肝臓学。60(2):576–87。土井:10.1002 /hep.26996。PMC 4083010。PMID 24492943。
^ Gao W、Xu Y、Liu J、Ho M。「Wntブロッキング抗体によるエピトープマッピング:ヘパラン硫酸のWnt結合ドメインの証拠」。ScientificReports。6:26245. Bibcode:2016NatSR … 626245G。土井:10.1038 / srep26245。PMC 4869111。PMID 27185050。
^ Gao W、Tang Z、Zhang YF、Feng M、Qian M、Dimitrov DS、Ho M。「グリピカン3を標的とする免疫毒素は、Wntシグナル伝達とタンパク質合成の二重阻害を介して肝臓癌を退行させます」。ネイチャーコミュニケーションズ。6:6536. Bibcode:2015NatCo … 6.6536G。土井:10.1038 / ncomms7536。PMC 4357278。PMID 25758784。
^ Li N、Wei L、Liu X、Bai H、Ye Y、Li D、他 。「グリピカン3の縮れたようなシステインに富むドメインは、Wnt結合を媒介し、マウスの肝細胞癌腫瘍の成長を調節します」。肝臓学。70(4):1231–1245。土井:10.1002 /hep.30646。PMC 6783318。PMID 30963603。
^ Ho M、Kim H。「グリピカン3:癌免疫療法の新しい標的」。European Journal ofCancer。47(3):333–8。土井:10.1016 /j.ejca.2010.10.024。PMC 3031711。PMID 21112773。
^ Li N、Gao W、Zhang YF、Ho M。「癌治療標的としてのグリピカン」。がんの動向。4(11):741–754。土井:10.1016 /j.trecan.2018.09.004。PMC 6209326。PMID 30352677。
^ 後主、魏; Xu、Yongmei; 劉、建; ホー、ミッチェル(2016年5月17日)。「Wntブロッキング抗体によるエピトープマッピング:ヘパラン硫酸のWnt結合ドメインの証拠」。ScientificReports。6:26245. Bibcode:2016NatSR … 626245G。土井:10.1038 / srep26245。ISSN 2045年から2322年。PMC 4869111。PMID 27185050。
^ Kolluri A、Ho M(2019-08-02)。「肝臓癌におけるWnt、YAP、およびヘッジホッグの調節におけるグリピカン-3の役割」。腫瘍学のフロンティア。9:708 DOI:10.3389 / fonc.2019.00708。PMC 6688162。PMID 31428581。
^ Malinauskas T、Aricescu AR、Lu W、Siebold C、Jones EY。「Wnt阻害因子1によるWntシグナル伝達阻害のモジュラーメカニズム」。自然構造および分子生物学。18(8):886–93。土井:10.1038 /nsmb.2081。PMC 3430870。PMID 21743455。
^ Malinauskas T。「ヒトWnt阻害因子-1のWIFドメインへの脂肪酸のドッキング」。脂質。43(3):227–30。土井:10.1007 / s11745-007-3144-3。PMID 18256869。S2CID 31357937。
^ Minde DP、Radli M、Forneris F、Maurice MM、RüdigerSG(2013)。「腺腫性ポリポーシスコリの大規模な障害は、点突然変異からWntシグナル伝達を保護する戦略を提供します」。PLOSONE。8(10):e77257。Bibcode:2013PLoSO … 877257M。土井:10.1371 /journal.pone.0077257。PMC 3793970。PMID 24130866。
^ ギルバートSF(2010)。発生生物学(第9版)。マサチューセッツ州サンダーランド:Sinauer Associates ISBN
9780878933846。
^ Vasiev B、Balter A、Chaplain M、Glazier JA、Weijer CJ。「ニワトリ胚の原腸陥入のモデル化:原始線条の形成」。PLOSONE。5(5):e10571。Bibcode:2010PLoSO … 510571V。土井:10.1371 /journal.pone.0010571。PMC 2868022。PMID 20485500。
^ ギルバートSF(2014)。「鳥の初期の発達」。発生生物学(第10版)。サンダーランド(MA):シナウアーアソシエイツ。
^ Ulloa F、MartíE。「Wntは戦争に勝った:Shhに対するWntの拮抗的役割は、脊椎動物の神経管の背腹パターン形成を制御する」。発生ダイナミクス。239(1):69–76。土井:10.1002 /dvdy.22058。PMID 19681160。
^ Zou Y。「軸索ガイダンスにおけるWntシグナル伝達」。神経科学の動向。27(9):528–32。土井:10.1016 /j.tins.2004.06.015。PMID 15331234。S2CID 15635026。
^ Gordon NK、Gordon R。「胚における分化の細胞小器官:細胞状態スプリッター」。理論生物学と医療モデリング。13:11。DOI:10.1186 / s12976-016-0037-2。PMC 4785624。PMID 26965444。
^ ゴードンN、ゴードン、R(2016)。胚発生の説明。シンガポール:世界科学出版。pp。580–591。土井:10.1142 / 8152。ISBN
978-981-4740-69-2。
^ Nusse R。「Wntシグナル伝達と幹細胞制御」。セルリサーチ。18(5):523–7。土井:10.1038 /cr.2008.47。PMID 18392048。
^ Bakre MM、Hoi A、Mong JC、Koh YY、Wong KY、Stanton LW。「持続的なWnt経路活性化を介したマウス胚性幹細胞からの多能性中内胚葉前駆細胞の生成」。Journal of BiologicalChemistry。282(43):31703–12。土井:10.1074 /jbc.M704287200。PMID 17711862。
^ Woll PS、Morris JK、Painschab MS、Marcus RK、Kohn AD、Biechele TL、Moon RT、Kaufman DS。「Wntシグナル伝達は、ヒト胚性幹細胞からの血液内皮細胞の発達を促進します」。血。111(1):122–31。土井:10.1182 / blood-2007-04-084186。PMC 2200802。PMID 17875805。
^ Schneider VA、Mercola M。「Wnt拮抗作用はアフリカツメガエルの心臓形成を開始します」。遺伝子と開発。15(3):304–15。土井:10.1101 /gad.855601。PMC 312618。PMID 11159911。
^ Marvin MJ、Di Rocco G、Gardiner A、Bush SM、Lassar AB。「Wnt活性の阻害は後部中胚葉からの心臓形成を誘発します」。遺伝子と開発。15(3):316–27。土井:10.1101 /gad.855501。PMC 312622。PMID 11159912。
^ Ueno S、Weidinger G、Osugi T、Kohn AD、Golob JL、Pabon L、Reinecke H、Moon RT、Murry CE。「ゼブラフィッシュおよび胚性幹細胞の心臓の仕様におけるWnt /β-カテニンシグナル伝達の二相性の役割」。アメリカ合衆国科学アカデミー紀要。104(23):9685–90。Bibcode:2007PNAS..104.9685U。土井:10.1073 /pnas.0702859104。PMC 1876428。PMID 17522258。
^ Willems E、Spiering S、Davidovics H、Lanier M、Xia Z、Dawson M、Cashman J、Mercola M。「Wnt経路の小分子阻害剤は、ヒト胚性幹細胞由来の中胚葉からの心筋細胞を強力に促進します」。循環研究。109(4):360–4。土井:10.1161 /CIRCRESAHA.111.249540。PMC 3327303。PMID 21737789。
^ Burridge PW、Matsa E、Shukla P、Lin ZC、Churko JM、Ebert AD、Lan F、Diecke S、Huber B、Mordwinkin NM、Plews JR、Abilez OJ、Cui B、Gold JD、Wu JC。「化学的に定義されたヒト心筋細胞の生成」。ネイチャーメソッズ。11(8):855–60。土井:10.1038 /nmeth.2999。PMC 4169698。PMID 24930130。
^ Kaldis P、Pagano M。「有糸分裂におけるWntシグナル伝達」。発生細胞。17(6):749–50。土井:10.1016 /j.devcel.2009.12.001。PMID 20059944。
^ Willert K、Jones KA。「Wntシグナル伝達:パーティーは核心にあるのか?」。遺伝子と開発。20(11):1394–404。土井:10.1101 /gad.1424006。PMID 16751178。
^ ホッジ、ラス(2016-01-25)。「癌幹細胞のプログラムをハッキングする」。Medicalxpress.com。メディカルエクスプレス。
^ シャンボニーA、ウェドリッチD(2013)。Wntシグナル伝達と細胞移動。マダムキュリーバイオサイエンスデータベース。ランデスバイオサイエンス。検索された5月7 2013。
^ Micalizzi DS、Farabough SM、Ford HL。「癌における上皮間葉転換:正常な発達と腫瘍の進行との類似点」。Journal of Mammary Gland Biology andNeoplasia。15(2):117–34。土井:10.1007 / s10911-010-9178-9。PMC 2886089。PMID 20490631。
^ Abiola M、Favier M、Christodoulou-Vafeiadou E、Pichard AL、Martelly I、Guillet-Deniau I。「Wnt /β-カテニンシグナル伝達の活性化は、骨格筋細胞におけるWnt10bとSREBP-1cの相互調節を通じてインスリン感受性を増加させます」。PLOSONE。4(12):e8509。Bibcode:2009PLoSO … 4.8509A。土井:10.1371 /journal.pone.0008509。PMC 2794543。PMID 20041157。
^ Milosevic、V。etal。Wnt /IL-1β/ IL-8オートクリン回路は、ABCB5.Intを誘導することにより、中皮腫開始細胞の化学療法抵抗性を制御します。J. Cancer、 https://doi.org/10.1002/ijc.32419
^ Howe LR、Brown AM。「Wntシグナル伝達と乳がん」。がんの生物学と治療。3(1):36–41。土井:10.4161 /cbt.3.1.561。PMID 14739782。
^ Taketo MM。「Wntシグナル活性化癌のシャットダウン」。ネイチャージェネティクス。36(4):320–2。土井:10.1038 / ng0404-320。PMID 15054482。
^ DiMeo TA、Anderson K、Phadke P、Fan C、Feng C、Perou CM、Naber S、Kuperwasser C。「新しい肺転移の特徴は、Wntシグナル伝達を、基底細胞様乳がんにおけるがん細胞の自己複製および上皮間葉転換と結び付けます」。がん研究。69(13):5364–73。土井:10.1158 /0008-5472.CAN-08-4135。PMC 2782448。PMID 19549913。
^ Anastas JN、Moon RT。「癌の治療標的としてのWNTシグナル伝達経路」。ネイチャーレビュー。がん。13(1):11–26。土井:10.1038 / nrc3419。PMID 23258168。S2CID 35599667。
^ Welters HJ、Kulkarni RN。「Wntシグナル伝達:ベータ細胞生物学および糖尿病との関連性」。内分泌学と代謝の傾向。19(10):349–55。土井:10.1016 /j.tem.2008.08.004。PMID 18926717。S2CID 19299033。
^ Yoon JC、Ng A、Kim BH、Bianco A、Xavier RJ、Elledge SJ。「Wntシグナル伝達はミトコンドリアの生理機能とインスリン感受性を調節します」。遺伝子と開発。24(14):1507–18。土井:10.1101 /gad.1924910。PMC 2904941。PMID 20634317。
^ Zhai L、Ballinger SW、Messina JL。「傷害誘発性インスリン抵抗性における活性酸素種の役割」。分子内分泌学。25(3):492–502。土井:10.1210 /me.2010-0224。PMC 3045736。PMID 21239612。
^ Grant SF、Thorleifsson G、Reynisdottir I、Benediktsson R、Manolescu A、Sainz J、他 。「転写因子7様2(TCF7L2)遺伝子の変異体は、2型糖尿病のリスクをもたらします」。ネイチャージェネティクス。38(3):320–3。土井:10.1038 / ng1732。PMID 16415884。S2CID 28825825。
参考文献
ミロシェビッチVら 。「Wnt /IL-1β/ IL-8オートクリン回路は、ABCB5を誘導することにより、中皮腫開始細胞の化学療法抵抗性を制御します」。Int。J.がん。146(1):192–207。土井:10.1002 /ijc.32419。PMID 31107974。S2CID 160014053。
Dinasarapu AR、Saunders B、Ozerlat I、Azam K、Subramaniam S。「シグナリングゲートウェイ分子ページ-データモデルの観点」。バイオインフォマティクス。27(12):1736–8。土井:10.1093 / bioinformatics / btr190。PMC 3106186。PMID 21505029。
外部リンク
米国国立医学図書館のWnt + Proteins Medical Subject Headings(MeSH)”