Z-DNA
Z-DNAは、多くの可能なの一つであり、二重らせん状の構造DNA。これは左巻きの二重らせん構造であり、より一般的なB-DNA形式のように、らせんが右ではなく左にジグザグパターンで巻かれています。Z-DNAは、A-DNAおよびB-DNAとともに、3つの生物学的に活性な二重らせん構造の1つであると考えられています。
Z-DNA構造。
プロテオペディア Z-DNA
コンテンツ
1 歴史
2 構造
2.1 Z-DNA構造の予測 2.2 B-DNAからのZ-DNAの形成経路
3 生物学的意義
3.1 転写におけるZ-DNA 3.2 Zαドメインの発見 3.3 ワクシニアE3Lタンパク質へのZ-DNA結合の結果
4 いくつかのDNAフォームの比較ジオメトリ
5 も参照してください
6 参考文献
歴史
左利きのDNAは、最初にによって発見されたロバート・ウェルズ繰り返しの彼らの研究中に、そして同僚ポリマーのイノシン-シトシン。彼らは、そのようなDNAの「逆」円二色性スペクトルを観察し、これを(正しく)解釈して、ストランドが左巻きで互いに巻き付いていることを意味しました。Z-DNAとより身近なB-DNAの関係は、PohlとJovinの研究によって示されました。彼は、ポリ(dG-dC)の紫外線円二色性が4M 塩化ナトリウムでほぼ反転することを示しました。解決。これがB-DNAからZ-DNAへの変換の結果であるという疑いは、これらの溶液とZ-DNA結晶のラマンスペクトルを調べることによって確認されました。その後、「Z-DNA」の結晶構造が発表され、DNA断片(自己相補的DNAヘキサマーd(CG)3)の最初の単結晶X線構造であることが判明しました。これは、ワトソン-クリックの塩基対によって結合された2つの逆平行鎖を持つ左巻きの二重らせんとして解決されました(X線結晶学を参照)。それは、1979年にMITでAndrew HJ Wang、Alexander Rich、および同僚によって解決されました。 2005年のB-to-Z-DNA接合部の結晶化は、細胞内でZ-DNAが果たす潜在的な役割についてのより良い理解を提供しました。Z-DNAのセグメントが形成されるときはいつでも、その両端にB–Z接合があり、ゲノムの残りの部分にあるB型のDNAに接続している必要が
2007年、Z-DNAのRNAバージョンであるZ- RNAは、A-RNA二重らせんを左巻きのらせんに変換したものとして説明されました。しかしながら、A-RNAからZ-RNAへの移行は1984年にすでに説明されています。
構造

Z-DNA結合ドメインに結合したB–Zジャンクション。2つの強調表示された押し出しベースに注意してPDBから :2ACJ。 Z-DNAは右利きの形式とはかなり異なります。実際、主な違いを説明するために、Z-DNAはB-DNAと比較されることがよくZ-DNAヘリックスは左巻きで、1つおきの塩基対を繰り返す構造になっています。A-およびB-DNAとは異なり、主溝と副溝は幅にほとんど違いがありません。この構造の形成は一般的に好ましくありませんが、特定の条件がそれを促進する可能性が交互のプリン-ピリミジン配列(特にポリ(dGC)2)、負のDNAスーパーコイルまたは高塩およびいくつかの陽イオン(すべて生理学的温度、37°C、およびpH 7.3–7.4)など。Z-DNAは、塩基対の押し出しを伴う構造でB-DNAとの接合部(「B-to-Z接合部ボックス」と呼ばれる)を形成できます。 Z-DNAコンフォメーションは、二重らせんの安定した特徴として存在しないため、研究が困難でした。代わりに、それは生物学的活動によって時折誘発され、その後すぐに消える一過性の構造です。
Z-DNA構造の予測
DNA配列がZ-DNA構造を形成する可能性を予測することが可能です。DNAがB型からZ型に反転する傾向を予測するためのアルゴリズムであるZHuntは、1984年にMITでP. ShingHoによって作成されました。このアルゴリズムは、後により開発されたトレーシー・キャンプ、P.クリストフチャンプ、サンドールモーリス、およびジェフリーM. Vargason(主任研究員としてホー有する)Z-DNAのゲノムワイドなマッピングのため。
B-DNAからのZ-DNAの形成経路
1979年のZ-DNAの発見と結晶化以来、この構成により、科学者はB-DNA構成からZ-DNA構成への経路とメカニズムについて戸惑いました。 B-DNAからZ-DNA構造へのコンフォメーション変化は原子レベルでは不明でしたが、2010年に、Lee etal。によってコンピューターシミュレーションが実施されました。BからZへの遷移の段階的な伝播が以前に仮定された協調メカニズムよりも低いエネルギー障壁を提供することを計算で決定することができました。これは計算によって証明されたので、さらなる確認と妥当性のために、経路を実験室で実験的にテストする必要が具体的には、彼らのジャーナル記事で、「現在の結果は、将来、単一分子FRET(smFRET)実験によってテストされる可能性がある」と述べています。 2018年、B-DNAからZ-DNAへの経路は、smFRETアッセイを使用して実験的に証明されました。これは、ドナーとアクセプターの蛍光色素(フルオロフォアとしても知られている)の間の強度値を、DNA分子にタグ付けされた状態で電子を交換する際の相互の関係で測定することによって実行されました。 フルオロフォア間の距離を使用して、色素の近接の変化とDNAのコンフォメーション変化を定量的に計算できます。Z-DNA高親和性結合タンパク質hZαADAR1 をさまざまな濃度で使用して、B-DNAからZ-DNAへの変換を誘導しました。 smFRETアッセイは、B-DNA構造に蓄積されたhZαADAR1の結合として形成され、それを安定化させたB *遷移状態を明らかにしました。このステップは、エネルギーの大きな破壊的な変化なしに、B-DNA構造がZ-DNA構造へのコンフォメーション変化を受けることを可能にする高い接合エネルギーを回避するために発生します。この結果は、Leeらの計算結果と一致しています。メカニズムが段階的であることを証明し、その目的は、B-DNAからZ-DNA構成へのコンフォメーション変化に対してより低いエネルギー障壁を提供することです。以前の概念とは異なり、結合タンパク質は、Z-DNAコンフォメーションが形成された後、実際には安定化されませんが、代わりに、B *コンフォメーションから直接Z-DNAの形成を促進します。高親和性タンパク質によって結合されているB-DNA構造。
生物学的意義
I型インターフェロン応答の調節におけるZ-DNAの生物学的役割は、遺伝性対側性ジストニア症(OMIM:127400)、エカルディ・グティエール症候群(OMIM:615010)、両側性線条体の3つのよく特徴付けられたまれなメンデルの法則の研究で確認されています。壊死/ジストニア。半数体ADARトランスクリプトームを持つ家族は、Zα変異体を直接疾患にマッピングすることを可能にし、遺伝情報が形状と配列の両方によってDNAにコード化されていることを示しています。がんにおけるI型インターフェロン応答の調節における役割は、腫瘍のパネルの40%が生存のためにADAR酵素に依存していたという発見によっても裏付けられています。
以前の研究では、Z-DNAはアルツハイマー病と全身性エリテマトーデスの両方に関連していた。これを示すために、正常で、アルツハイマー病に中程度の影響を受け、アルツハイマー病に重度の影響を受けた脳の海馬で見つかったDNAについて研究が行われました。円二色性を使用することにより、この研究は、深刻な影響を受けた人々のDNAにZ-DNAが存在することを示しました。この研究では、中程度に影響を受けたDNAの大部分がBZ中間コンフォメーションにあることもわかった。これらの発見から、B-DNAからZ-DNAへの移行はアルツハイマー病の進行に依存していると結論付けられたため、これは重要です。さらに、Z-DNAは、天然に存在する抗体の存在を介して全身性エリテマトーデス(SLE)と関連しています。かなりの量の抗Z-DNA抗体がSLE患者に見られ、他のリウマチ性疾患には存在しませんでした。これらの抗体には2つのタイプがラジオイムノアッセイにより、一方はZ-DNAおよび変性DNAの表面に露出した塩基と相互作用し、もう一方はZ-DNAのみのジグザグバックボーンとのみ相互作用することがわかりました。アルツハイマー病に見られるものと同様に、抗体は疾患の病期によって異なり、SLEの最も活発な病期に最大の抗体が
転写におけるZ-DNA
Z-DNAは、転写中にねじれひずみを緩和すると一般に考えられており、負のスーパーコイルに関連しています。 ただし、スーパーコイルはDNAの転写と複製の両方に関連していますが、Z-DNAの形成は主に転写の速度に関連しています。
ヒト22番染色体の研究は、核内因子IのZ-DNA形成領域とプロモーター領域の間に相関関係があることを示しました。これは、いくつかのヒト遺伝子の転写がZ-DNA形成と核内因子Iの活性化によって調節されている可能性があることを示唆しています。
プロモーター領域の下流のZ-DNA配列は、転写を刺激することが示されています。活性の最大の増加は、Z-DNA配列がプロモーター配列の3ヘリックスターン後に配置されたときに観察されます。さらに、Z-DNAがヌクレオソームを形成する可能性は低く、ヌクレオソームはZ-DNA形成配列の後に位置することがよくこの特性のため、Z-DNAはヌクレオソームポジショニングをコードすると仮定されています。ヌクレオソームの配置は転写因子の結合に影響を与えるため、Z-DNAは転写速度を調節すると考えられています。
負のスーパーコイルを介したRNAポリメラーゼの経路の背後で開発された、アクティブな転写を介して形成されたZ-DNAは、遺伝子の不安定性を高め、プロモーターの近くで突然変異誘発の傾向を生み出すことが示されています。 Escherichia coliに関する研究では、Z-DNA形成配列を含むプラスミド領域で遺伝子の欠失が自発的に発生することがわかりました。哺乳類細胞では、そのような配列の存在は、染色体の二本鎖切断のために大きなゲノム断片の欠失を生み出すことが見出された。腫瘍細胞の破壊領域がZ-DNA形成配列の周りにプロットされているため、これらの遺伝子改変は両方とも、白血病やリンパ腫などの癌に見られる遺伝子転座に関連しています。しかしながら、細菌プラスミドの小さな欠失は複製のずれに関連しているが、哺乳類細胞に関連する大きな欠失は、エラーを起こしやすいことが知られている非相同末端結合修復によって引き起こされている。
トリパノソーマに対する臭化エチジウム(EtBr)の毒性作用は、キネトプラスト類のDNAがZ型にシフトすることによって引き起こされます。シフトは、EtBrのインターカレーションと、それに続くDNA構造の緩みによって引き起こされ、DNAの巻き戻し、Z型へのシフト、およびDNA複製の阻害につながります。
Zαドメインの発見
Z-DNAに高い親和性で結合する最初のドメインは、AlanHerbertによって開発されたアプローチを使用してADAR1で発見されました。 結晶学的およびNMR研究により、このドメインが非配列特異的にZ-DNAに結合するという生化学的所見が確認された。 関連するドメインは、配列相同性を介して他の多くのタンパク質で同定されました。 Zαドメインの同定は、Z-RNAおよびB-Z接合部の特性評価につながる他の結晶学的研究のためのツールを提供しました。生物学的研究は、ADAR1のZ-DNA結合ドメインが、新しく形成されたRNAの配列を活発な転写部位に修飾するこの酵素を局在化させる可能性があることを示唆しました。 ヒトにおけるAluレトロエレメントの侵入に対するゲノムの防御におけるZα、Z-DNAおよびZ-RNAの役割は、dsRNAに対する自然免疫応答の調節のメカニズムに進化しました。Zαの変異は、メンデルの法則エカルディ・グティエール症候群などのヒトの干渉障害の原因です。
ワクシニアE3Lタンパク質へのZ-DNA結合の結果
Z-DNAがより徹底的に研究されるにつれて、Z-DNAの構造がロンドン分散と水素結合を介してZ-DNA結合タンパク質に結合できることが発見されました。 Z-DNA結合タンパク質の一例はワクシニアE3Lタンパク質であり、これはE3L遺伝子の産物であり、Z-DNAに結合する哺乳類タンパク質を模倣している。 E3Lタンパク質はZ-DNAに親和性があるだけでなく、ポックスウイルスの一種であるワクシニアウイルスによって引き起こされるマウスの病原性の重症度のレベルにも関与していることがわかっています。病原性を決定するE3Lタンパク質の2つの重要な要素は、N末端とC末端です。N末端はZαドメインと同様の配列で構成されており、アデノシンデアミナーゼz-αドメインとも呼ばれますが、C末端は二本鎖RNA結合モチーフで構成されています。キム、Y。らによって行われた研究を通して。マサチューセッツ工科大学では、E3Lタンパク質のN末端をE3Lと同様の14個のZ-DNA結合残基を含むZαドメイン配列で置き換えると、マウスのウイルスの病原性にほとんどまたはまったく影響がないことが示されました。対照的に、Kim、Y。etal。また、E3LのN末端の83残基すべてを削除すると、病原性が低下することもわかりました。これは、Z-DNA結合残基を含むN末端が病原性に必要であるという彼らの主張を裏付けています。全体として、これらの発見は、E3Lタンパク質のN末端内の同様のZ-DNA結合残基とZαドメインがワクシニアウイルスによって引き起こされる毒性を決定する最も重要な構造因子であり、アミノ酸残基は関与していないことを示していますZ-DNA結合は、ほとんどまたはまったく効果がありません。これらの発見の将来の意味は、ワクシニアウイルスを含むワクチンにおけるE3LのZ-DNA結合を減少させることを含み、それによりウイルスに対する陰性反応をヒトで最小限に抑えることができます。
さらに、AlexanderRichとJin-AhKwonは、E3LがヒトIL-6、NF-AT、およびp53遺伝子のトランス活性化因子として機能することを発見しました。彼らの結果は、E3Lを含むHeLa細胞がヒトIL-6、NF-AT、およびp53遺伝子の発現を増加させ、特定のZ-DNA結合アミノ酸残基の点突然変異または欠失がその発現を減少させたことを示しています。具体的には、Tyr48およびPro63の突然変異は、E3LとZ-DNAの間の水素結合およびロンドン分散力の喪失の結果として、前述の遺伝子のトランス活性化を低下させることがわかった。全体として、これらの結果は、Z-DNAとZ-DNA結合タンパク質間の結合と相互作用を減少させると、毒性と遺伝子発現の両方が減少することを示し、したがって、Z-DNAとE3L結合タンパク質の間に結合を持つことの重要性を示しています。
いくつかのDNAフォームの比較ジオメトリ

A-、B-、およびZ-DNAの側面図。
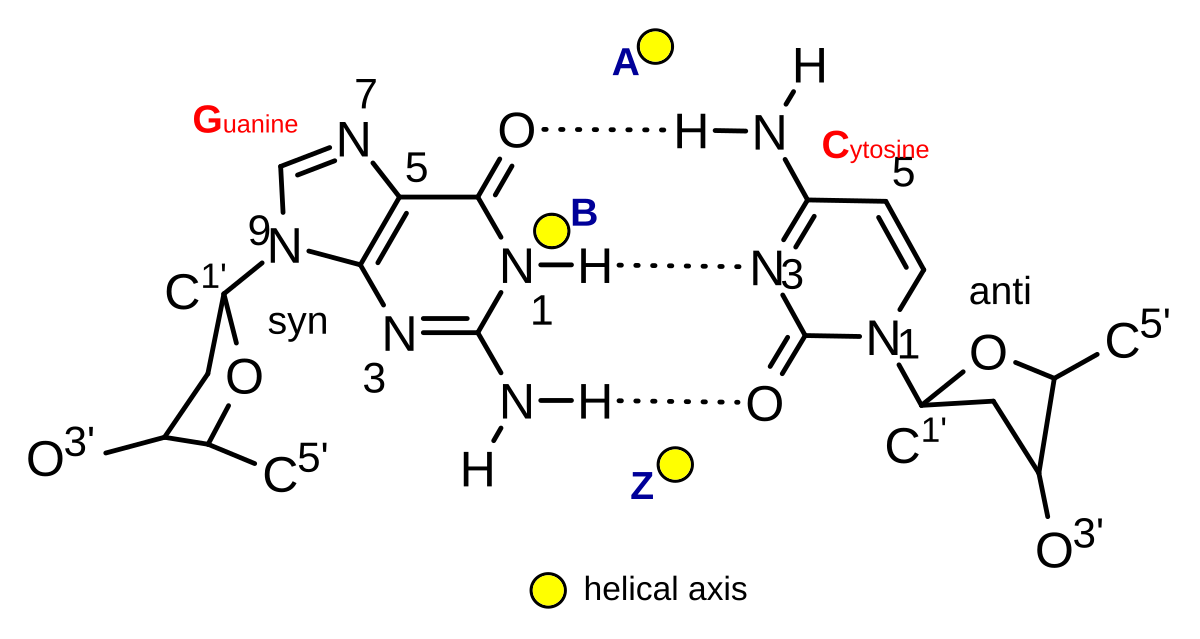
A-、B-、およびZ-DNAのらせん軸。
A-、B、およびZ-DNAのジオメトリ属性
Aフォーム Bフォーム Zフォーム
らせん感覚 右利き 右利き 左利き
繰り返し単位1 bp 1 bp 2 bp
回転/ bp
32.7°
34.3° 30° bp /ターン11 10 12
軸に対するbpの傾斜+ 19°
−1.2°
−9°
軸に沿った上昇/ bp
2.3Å(0.23 nm)
3.32Å(0.332 nm)
3.8Å(0.38 nm)
らせんのピッチ/ターン
28.2Å(2.82 nm)
33.2Å(3.32 nm)
45.6Å(4.56 nm)
平均プロペラねじれ+ 18°+ 16° 0° グリコシル角 アンチ アンチ
C:アンチ、G:シン
シュガーパッカー
C3′-エンド
C2′-エンド
C:C2′-エンド、G:C3′-エンド
直径
23Å(2.3 nm)
20Å(2.0 nm)
18Å(1.8 nm)
も参照してください ADAR1 DNAスーパーコイル E3L DNAの機械的性質
プロテオペディア Z-DNA
サテライトDNA
Z-DNA結合タンパク質1(ZBP1)
ズオティン
参考文献
^ 三井康夫; ラングリッジ、R。; ショートル、BE; カンター、CR; グラント、RC; 児玉眞; ウェルズ、RD(1970)。「珍しい二重らせんDNAであるポリd(I–C)・ポリd(I–C)の物理的および酵素的研究」。自然。228(5277):1166–1169。土井:10.1038 / 2281166a0。PMID 4321098。
^ ポール、FM; Jovin、TM(1972)。「合成DNAの塩による協同的コンフォメーション変化:ポリ(dG-dC)による平衡および速度論的研究」。分子生物学ジャーナル。67(3):375–396。土井:10.1016 / 0022-2836(72)90457-3。PMID 5045303。
^ Thamann、TJ; 主、RC; 王、AH; リッチ、A。(1981)。「ポリ(dG–dC)・ポリ(dG–dC)の高塩形態は左利きのZ-DNA:結晶と溶液のラマンスペクトル」。核酸研究。9(20):5443–5457。土井:10.1093 / nar /9.20.5443。PMC 327531。PMID 7301594。
^ 王、AH; Quigley、GJ; コルパック、FJ; クロフォード、JL; ヴァンブーム、JH; van der Marel、G。; リッチ、A。(1979)。「原子分解能での左巻き二重らせんDNA断片の分子構造」。自然。282(5740):680–686。Bibcode:1979Natur.282..680W。土井:10.1038 / 282680a0。PMID 514347。
^ Ha、SC; Lowenhaupt、K。; リッチ、A。; キム、YG; キム株式会社(2005)。「B-DNAとZ-DNAの間の接合部の結晶構造は、2つの押し出された塩基を明らかにします」。自然。437(7062):1183–1186。Bibcode:2005Natur.437.1183H。土井:10.1038 / nature04088。PMID 16237447。
^ Placido、D。; ブラウン、BA、II; Lowenhaupt、K。; リッチ、A。; アサナシアディス、A。(2007)。「RNA編集酵素ADAR1のZalphaドメインによって結合された左巻きのRNA二重らせん」。構造。15(4):395–404。土井:10.1016 /j.str.2007.03.001。PMC 2082211。PMID 17437712。
^ ホール、K。; Cruz、P。; Tinoco、I.、Jr; Jovin、TM; van de Sande、JH(1984年10月)。「’ Z-RNA’—左巻きのRNA二重らせん」。自然。311(5986):584–586。Bibcode:1984Natur.311..584H。土井:10.1038 / 311584a0。PMID 6482970。
^ Rosa、M。; de Sanctis、D。; ロザリオ、AL; アーチャー、M。; リッチ、A。; Athanasiadis、A。; マサチューセッツ州カロンド。「2つのZ-DNAヘリックス間の接合部の結晶構造」。国立科学アカデミーの議事録。107(20):9088–9092。Bibcode:2010PNAS..107.9088D。土井:10.1073 /pnas.1003182107。PMC 2889044。PMID 20439751。
^ 張、H。; Yu、H。; レン、J。; Qu、X。(2006)。「低塩条件下での可逆的B / Z-DNA転移およびキュバン様ユーロピウム-L-アスパラギン酸複合体による非B型ポリ(dA)ポリ(dT)選択性」。生物物理ジャーナル。90(9):3203–3207。Bibcode:2006BpJ …. 90.3203Z。土井:10.1529 /biophysj.105.078402。PMC 1432110。PMID 16473901。
^ ホー、PS; エリソン、MJ; Quigley、GJ; リッチ、A。(1986)。「自然に発生する配列におけるZ-DNAの形成を予測するためのコンピューター支援熱力学的アプローチ」。EMBOジャーナル。5(10):2737–2744。土井:10.1002 /j.1460-2075.1986.tb04558.x。PMC 1167176。PMID 3780676。
^ Champ、PC; モーリス、S。; バルガソン、JM; キャンプ、T。; ホー、PS(2004)。「ヒト22番染色体におけるZ-DNAと核内因子Iの分布:結合した転写調節のモデル」。核酸研究。32(22):6501–6510。土井:10.1093 / nar / gkh988。PMC 545456。PMID 15598822。
^ 王、アンドリューH.-J。; Quigley、Gary J。; Kolpak、Francis J。; クロフォード、ジェームズL。; ヴァンブーム、ジャックH。; van der Marel、Gijs; リッチ、アレクサンダー(1979年12月)。「原子分解能での左巻き二重らせんDNA断片の分子構造」。自然。282(5740):680–686。Bibcode:1979Natur.282..680W。土井:10.1038 / 282680a0。ISSN 0028から0836まで。PMID 514347。
^ 李周勇、キム、ヤンギュン; キム、キョンギュ; ソク、チャオク(2010-08-05)。「B-DNAとZ-DNAの間の移行:B-Zジャンクション伝播の自由エネルギー地形」。物理化学Bのジャーナル。114(30):9872–9881。CiteSeerX 10.1.1.610.1717。土井:10.1021 / jp103419t。ISSN 1520から6106まで。
^ キム、スクホー; リム、ソヒ; Lee、Ae-Ree; クォン・ドゥホーン; ソン、ヒョンキュ; Lee、Joon-Hwa; チョ、ミンヘン; ジョナー、アルバート; イ・ナムギョン(2018-03-23)。「タンパク質によって誘発されるB–Z遷移におけるZ-DNAへの経路を明らかにする」。核酸研究。46(8):4129–4137。土井:10.1093 / nar / gky200。ISSN 0305から1048まで。PMC 5934635。PMID 29584891。
^ クーパー、デビッド; うーん、ホイ; トーザン、ローレンスJ。; Poddar、Nitesh; Landes、Christy F.(2013-06-03)。「さまざまな光防護システムの存在下での単一分子フェルスター共鳴エネルギー移動に使用されるシアニンフルオロフォアの光退色寿命」。ChemBioChem。14(9):1075〜1080。土井:10.1002 /cbic.201300030。ISSN 1439から4227まで。PMC 3871170。PMID 23733413。
^ Didenko、Vladimir V.。「蛍光共鳴エネルギー移動(FRET)を使用したDNAプローブ:設計と応用」。バイオテクニック。31(5):1106–1121。土井:10.2144 / 01315rv02。ISSN 0736から6205まで。PMC 1941713。PMID 11730017。
^ ハーバート、A。; アルフケン、J。; キム、Y.-G。; ミアン、IS; 西倉健一; リッチ、A。(1997-08-05)。「ヒト編集酵素、二本鎖RNAアデノシンデアミナーゼに存在するZ-DNA結合ドメイン」。国立科学アカデミーの議事録。94(16):8421–8426。Bibcode:1997PNAS … 94.8421H。土井:10.1073 /pnas.94.16.8421。ISSN 0027から8424まで。PMC 22942。PMID 9237992。
^ ハーバート、A。(2019)。「二本鎖RNA編集酵素ADARのZαドメインによるZ-DNAおよびZ-RNAの認識に影響を与える変異体によって引き起こされるメンデルの法則」。ヒト遺伝学のヨーロッパジャーナル。8:114〜117。土井:10.1038 / s41431-019-0458-6。PMC 6906422。PMID 31320745。
^ ハーバート、A。(2019)。「癌におけるADARと免疫サイレンシング」。がんの動向。5(5):272–282。土井:10.1016 /j.trecan.2019.03.004。PMID 31174840。
^ Suram、Anitha; Rao、Jagannatha KS; S.、Latha K。; A.、Viswamitra M.(2002)。「アルツハイマー病の脳の海馬におけるB-DNAからZ-DNAコンフォメーションへのDNAのトポロジー変化を示す最初の証拠」。NeuroMolecularMedicine。2(3):289–298。土井:10.1385 / nmm:2:3:289。ISSN 1535年から1084年。
^ Lafer、EM; ヴァッレ、RP; Möller、A; ノードハイム、A; Schur、PH; リッチ、A; Stollar、BD(1983-02-01)。「ヒト全身性エリテマトーデスにおけるZ-DNA特異的抗体」。臨床調査ジャーナル。71(2):314–321。土井:10.1172 / jci110771。ISSN 0021から9738まで。PMC 436869。PMID 6822666。
^ リッチ、A; 張、S(2003)。「タイムライン:Z-DNA:生物学的機能への長い道のり」。ネイチャーレビュー遺伝学。4(7):566–572。土井:10.1038 / nrg1115。PMID 12838348。
^ Wittig、B。; ドービック、T。; リッチ、A。(1991)。「転写は、代謝的に活性な透過性の哺乳類細胞核におけるZ-DNA形成に関連しています」。国立科学アカデミーの議事録。88(6):2259–2263。Bibcode:1991PNAS … 88.2259W。土井:10.1073 /pnas.88.6.2259。PMC 51210。PMID 2006166。
^ ウォン、B。; Chen、S。; クォン、J.-A。; リッチ、A。(2007)。「酵母Saccharomycescerevisiaeのヌクレオソーム境界要素としてのZ-DNAの特性評価」。国立科学アカデミーの議事録。104(7):2229–2234。Bibcode:2007PNAS..104.2229W。土井:10.1073 /pnas.0611447104。PMC 1892989。PMID 17284586。
^ Wang、G。; クリステンセン、LA; バスケス、KM(2006)。「Z-DNA形成配列は哺乳類細胞に大規模な欠失を生成します」。国立科学アカデミーの議事録。108(8):2677–2682。Bibcode:2006PNAS..103.2677W。土井:10.1073 /pnas.0511084103。PMC 1413824。PMID 16473937。
^ フロイント、AM; ビチャラ、M。; フックス、RP(1989)。「Z-DNA形成配列は自発的な欠失のホットスポットです」。国立科学アカデミーの議事録。86(19):7465–7469。Bibcode:1989PNAS … 86.7465F。土井:10.1073 /pnas.86.19.7465。PMC 298085。PMID 2552445。
^ Roy Chowdhury、A。; バクシ、R。; 王、J。; Yıldırır、G。; 劉、B。; Pappas-Brown、V。; Tolun、G。; グリフィス、JD; シャピロ、TA; ジェンセン、RE; イングランド、PT。「エチジウムブロマイドによるアフリカトリパノソームの殺害」。PLoS病原体。6(12):e1001226。土井:10.1371 /journal.ppat.1001226。PMC 3002999。PMID 21187912。
^ ハーバート、A。; リッチ、A。(1993)。「線形オリゴデオキシヌクレオチドを使用してZ-DNA結合タンパク質を同定および特性評価する方法」。核酸研究。21(11):2669–2672。土井:10.1093 / nar /21.11.2669。PMC 309597。PMID 8332463。
^ ハーバート、A。; アルフケン、J。; キム、YG; ミアン、IS; 西倉健一; リッチ、A。(1997)。「ヒト編集酵素、二本鎖RNAアデノシンデアミナーゼに存在するZ-DNA結合ドメイン」。国立科学アカデミーの議事録。94(16):8421–8426。Bibcode:1997PNAS … 94.8421H。土井:10.1073 /pnas.94.16.8421。PMC 22942。PMID 9237992。
^ ハーバート、A。; シェード、M。; Lowenhaupt、K。; アルフケン、J; シュワルツ、T。; Shlyakhtenko、LS; リュブチェンコ、YL; リッチ、A。(1998)。「ヒトADAR1のZαドメインは、多くの異なる配列のZ-DNAコンフォーマーに結合します」。核酸研究。26(15):2669–2672。土井:10.1093 / nar /26.15.3486。PMC 147729。PMID 9671809。
^ Schwartz、T。; マサチューセッツ州ルルド; Lowenhaupt、K。; ハーバート、A。; リッチ、A。(1999)。「左巻きZ-DNAに結合したヒト編集酵素ADAR1のZαドメインの結晶構造」。科学。284(5421):1841–1845。土井:10.1126 /science.284.5421.1841。PMID 10364558。
^ Schade、M。; ターナー、CJ; Kühne、R。; Schmieder、P。; Lowenhaupt、K。; ハーバート、A。; リッチ、A。; Oschkinat、H(1999)。「ヒトRNA編集酵素ADAR1のZαドメインの溶液構造は、Z-DNAの事前配置された結合表面を明らかにします」。国立科学アカデミーの議事録。96(22):2465–2470。Bibcode:1999PNAS … 9612465S。土井:10.1073 /pnas.96.22.12465。PMC 22950。PMID 10535945。
^ ハーバート、A。; リッチ、A。(2001)。「におけるdsRNAおよびZ-DNAに対する結合ドメインの役割インビボADAR1によって編集最小の基板」。国立科学アカデミーの議事録。98(21):12132–12137。Bibcode:2001PNAS … 9812132H。土井:10.1073 /pnas.211419898。PMC 59780。PMID 11593027。
^ Halber、D。(1999-09-11)。「科学者は「左利きの」DNAの生物活性を観察します」。MITニュースオフィス。
^ ハーバート、A。(2019)。「人間の病気におけるZ-DNAとZ-RNA」。コミュニケーション生物学。2:7。DOI:10.1038 / s42003-018-0237-X 。PMC 6323056。PMID 30729177。
^ Kwon、J.-A。; リッチ、A。(2005-08-26)。「ワクシニアウイルスZ-DNA結合タンパク質E3Lの生物学的機能:HeLa細胞における遺伝子トランス活性化と抗アポトーシス活性」。国立科学アカデミーの議事録。102(36):12759–12764。土井:10.1073 /pnas.0506011102。ISSN 0027から8424まで。
^ Kim、Y.-G。; Muralinath、M。; Brandt、T。; パーシー、M。; Hauns、K。; Lowenhaupt、K。; ジェイコブス、BL; リッチ、A。(2003-05-30)。「ワクシニアウイルスの病因におけるZ-DNA結合の役割」。国立科学アカデミーの議事録。100(12):6974–6979。土井:10.1073 /pnas.0431131100。ISSN 0027から8424まで。PMC 165815。PMID 12777633。
^ キム、Y.-G。; Lowenhaupt、K。; ああ、D.-B。; キム、KK; リッチ、A。(2004-02-02)。「ワクシニア病原性因子E3LがinvivoでZ-DNAに結合するという証拠:ポックスウイルス感染症の治療法の開発への影響」。国立科学アカデミーの議事録。101(6):1514〜1518。土井:10.1073 /pnas.0308260100。ISSN 0027から8424まで。PMC 341766。PMID 14757814。
^ シンデン、リチャードR.(1994)。DNAの構造と機能(第1版)。アカデミックプレス。NS。398. ISBN
978-0-126-45750-6。
^ リッチ、A。; Norheim、A。; 王、AH(1984)。「左利きのZ-DNAの化学と生物学」。生化学の年次レビュー。53(1):791–846。土井:10.1146 /annurev.bi.53.070184.004043。PMID 6383204。
^ Ho、PS(1994-09-27)。「d(CA / TG)nの非B-DNA構造はZ-DNAのそれと異ならない」。国立科学アカデミーの議事録。91(20):9549–9553。Bibcode:1994PNAS … 91.9549H。土井:10.1073 /pnas.91.20.9549。PMC 44850。PMID 7937803。
“